

A new front in cell invasion: The invadopodial membrane
- PMID: 27402208
- PMCID: PMC5110387
- DOI: 10.1016/j.ejcb.2016.06.006
A new front in cell invasion: The invadopodial membrane
Abstract
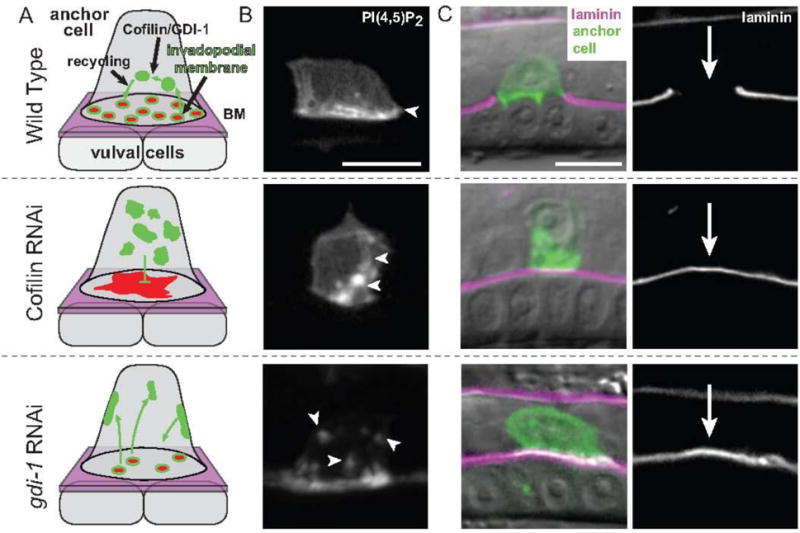
Invadopodia are F-actin-rich membrane protrusions that breach basement membrane barriers during cell invasion. Since their discovery more than 30 years ago, invadopodia have been extensively investigated in cancer cells in vitro, where great advances in understanding their composition, formation, cytoskeletal regulation, and control of the matrix metalloproteinase MT1-MMP trafficking have been made. In contrast, few studies examining invadopodia have been conducted in vivo, leaving their physiological regulation unclear. Recent live-cell imaging and gene perturbation studies in C. elegans have revealed that invadopodia are formed with a unique invadopodial membrane, defined by its specialized lipid and associated protein composition, which is rapidly recycled through the endolysosome. Here, we provide evidence that the invadopodial membrane is conserved and discuss its possible functions in traversing basement membrane barriers. Discovery and examination of the invadopodial membrane has important implications in understanding the regulation, assembly, and function of invadopodia in both normal and disease settings.
Keywords: Basement membrane; C. elegans; Cell invasion; Endocytic membrane trafficking; Invadopodia; Invadopodial membrane.
Copyright © 2016 Elsevier GmbH. All rights reserved.
Figures
Similar articles
-
ADF/cofilin promotes invadopodial membrane recycling during cell invasion in vivo.J Cell Biol. 2014 Mar 31;204(7):1209-18. doi: 10.1083/jcb.201312098. Epub 2014 Mar 24. J Cell Biol. 2014. PMID: 24662568 Free PMC article.
-
Matrix invasion by tumour cells: a focus on MT1-MMP trafficking to invadopodia.J Cell Sci. 2009 Sep 1;122(Pt 17):3015-24. doi: 10.1242/jcs.034561. J Cell Sci. 2009. PMID: 19692588 Review.
-
Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function.Cancer Res. 2006 Mar 15;66(6):3034-43. doi: 10.1158/0008-5472.CAN-05-2177. Cancer Res. 2006. PMID: 16540652
-
A Sensitized Screen for Genes Promoting Invadopodia Function In Vivo: CDC-42 and Rab GDI-1 Direct Distinct Aspects of Invadopodia Formation.PLoS Genet. 2016 Jan 14;12(1):e1005786. doi: 10.1371/journal.pgen.1005786. eCollection 2016 Jan. PLoS Genet. 2016. PMID: 26765257 Free PMC article.
-
Secretory and endo/exocytic trafficking in invadopodia formation: the MT1-MMP paradigm.Eur J Cell Biol. 2011 Feb-Mar;90(2-3):108-14. doi: 10.1016/j.ejcb.2010.04.007. Eur J Cell Biol. 2011. PMID: 20605060 Review.
Cited by
-
Tumor Cellular and Microenvironmental Cues Controlling Invadopodia Formation.Front Cell Dev Biol. 2020 Oct 15;8:584181. doi: 10.3389/fcell.2020.584181. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33178698 Free PMC article. Review.
-
Live-cell confocal microscopy and quantitative 4D image analysis of anchor-cell invasion through the basement membrane in Caenorhabditis elegans.Nat Protoc. 2017 Oct;12(10):2081-2096. doi: 10.1038/nprot.2017.093. Epub 2017 Sep 7. Nat Protoc. 2017. PMID: 28880279 Free PMC article.
-
Suppression of the Growth and Invasion of Human Head and Neck Squamous Cell Carcinomas via Regulating STAT3 Signaling and the miR-21/β-catenin Axis with HJC0152.Mol Cancer Ther. 2017 Apr;16(4):578-590. doi: 10.1158/1535-7163.MCT-16-0606. Epub 2017 Jan 30. Mol Cancer Ther. 2017. PMID: 28138036 Free PMC article.
-
Inhibition of ovarian tumor cell invasiveness by targeting SYK in the tyrosine kinase signaling pathway.Oncogene. 2018 Jul;37(28):3778-3789. doi: 10.1038/s41388-018-0241-0. Epub 2018 Apr 11. Oncogene. 2018. PMID: 29643476 Free PMC article.
-
Integrins: Moonlighting Proteins in Invadosome Formation.Cancers (Basel). 2019 May 2;11(5):615. doi: 10.3390/cancers11050615. Cancers (Basel). 2019. PMID: 31052560 Free PMC article. Review.
References
-
- Altincicek B, Fischer M, Luersen K, Boll M, Wenzel U, Vilcinskas A. Role of matrix metalloproteinase ZMP-2 in pathogen resistance and development in Caenorhabditis elegans. Dev Comp Immunol. 2010;34:1160–1169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
