

ChloroSeq, an Optimized Chloroplast RNA-Seq Bioinformatic Pipeline, Reveals Remodeling of the Organellar Transcriptome Under Heat Stress
- PMID: 27402360
- PMCID: PMC5015939
- DOI: 10.1534/g3.116.030783
ChloroSeq, an Optimized Chloroplast RNA-Seq Bioinformatic Pipeline, Reveals Remodeling of the Organellar Transcriptome Under Heat Stress
Abstract
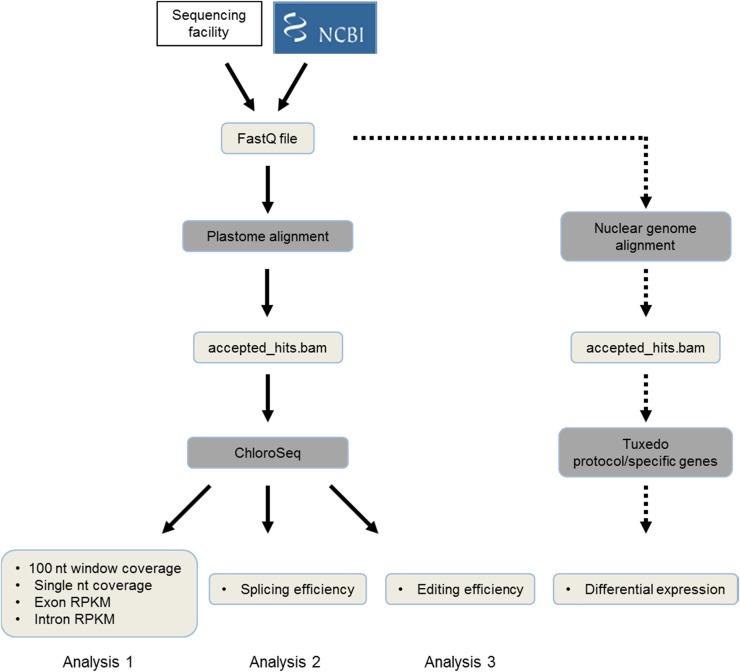
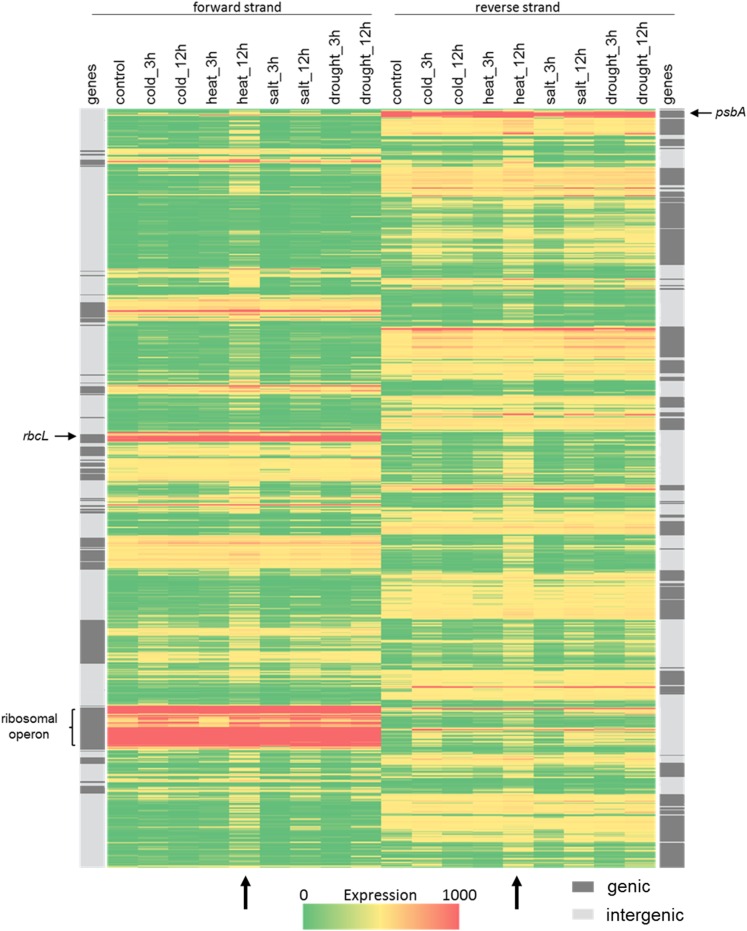
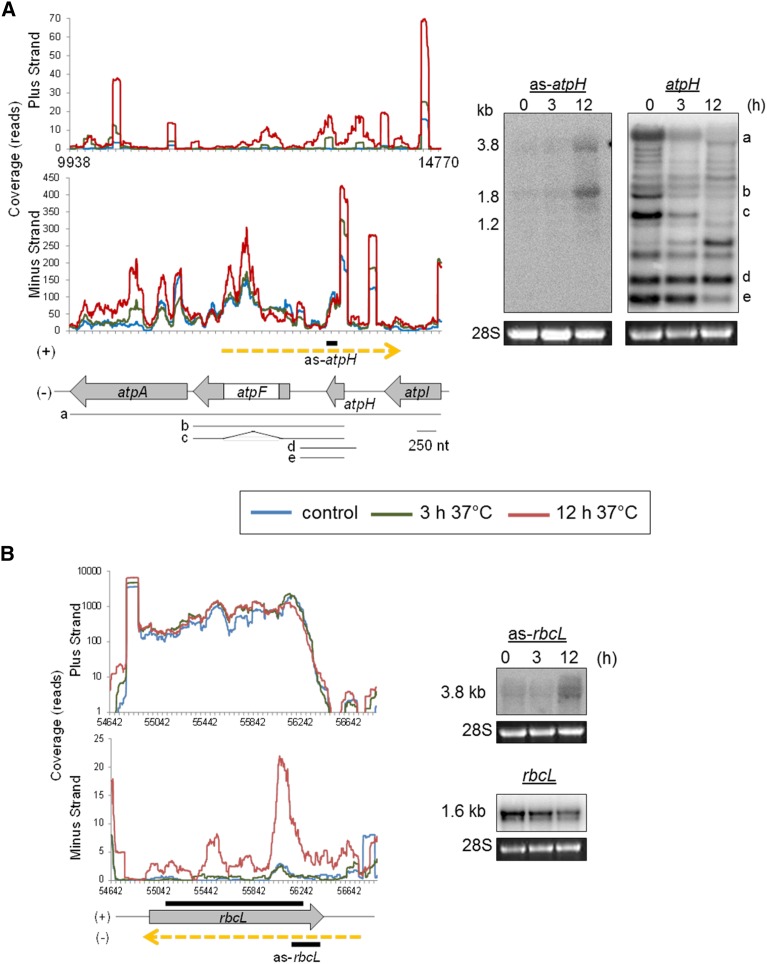
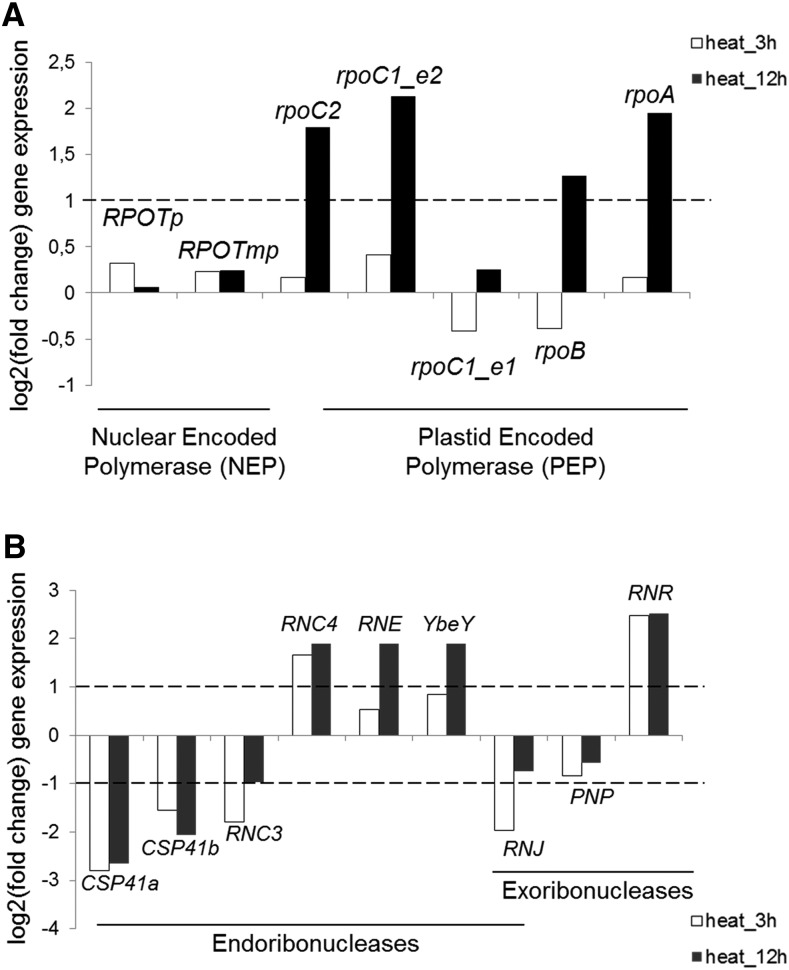
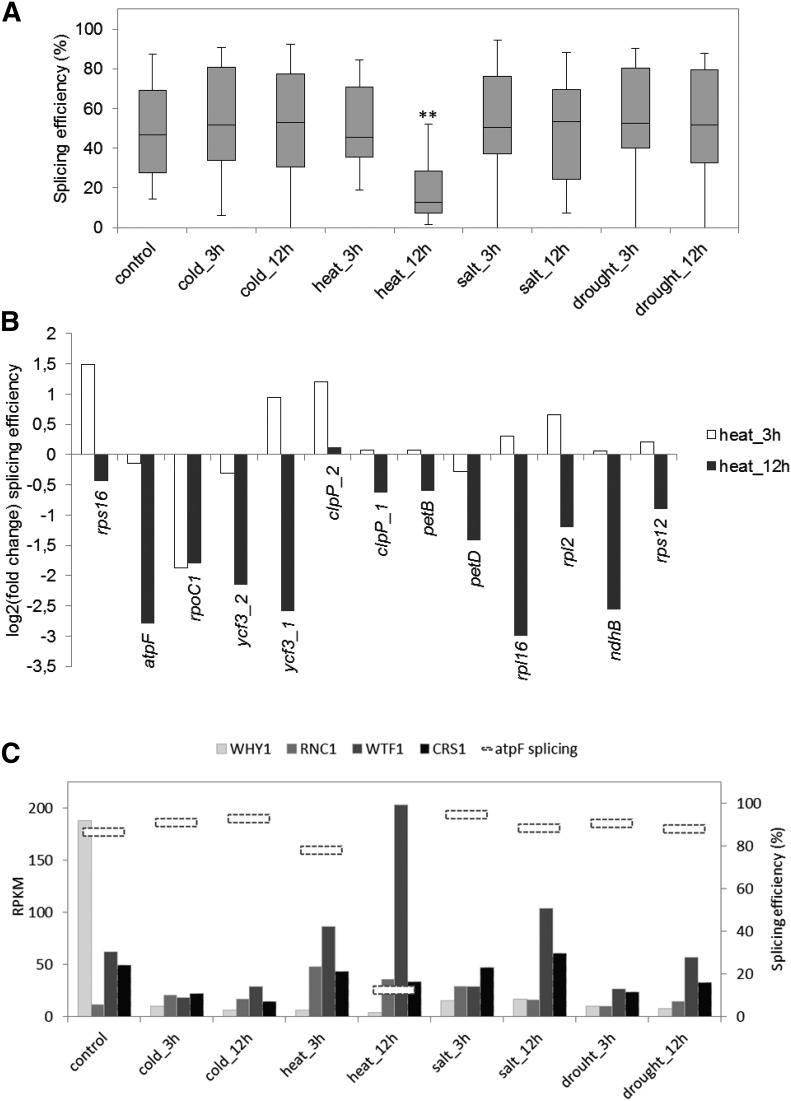
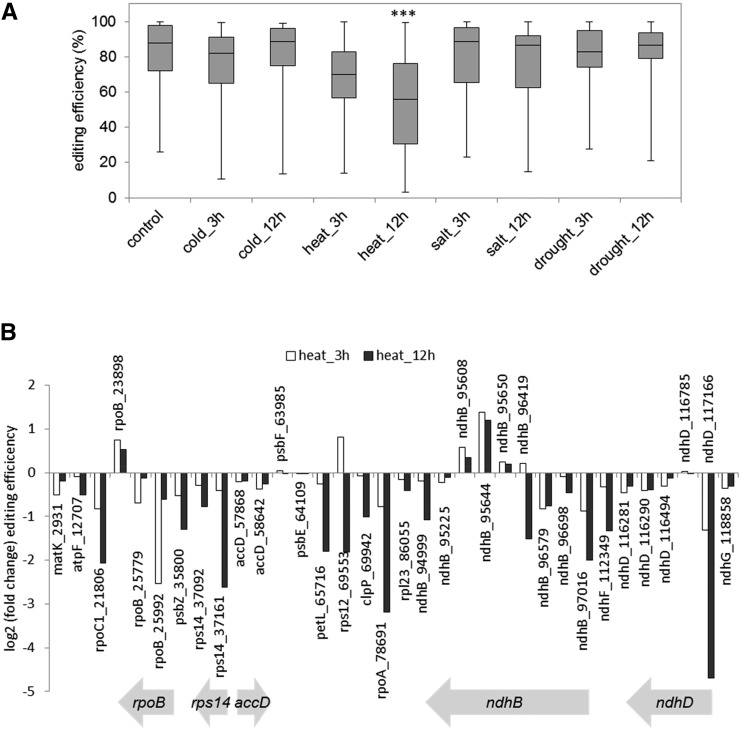
Although RNA-Seq has revolutionized transcript analysis, organellar transcriptomes are rarely assessed even when present in published datasets. Here, we describe the development and application of a rapid and convenient method, ChloroSeq, to delineate qualitative and quantitative features of chloroplast RNA metabolism from strand-specific RNA-Seq datasets, including processing, editing, splicing, and relative transcript abundance. The use of a single experiment to analyze systematically chloroplast transcript maturation and abundance is of particular interest due to frequent pleiotropic effects observed in mutants that affect chloroplast gene expression and/or photosynthesis. To illustrate its utility, ChloroSeq was applied to published RNA-Seq datasets derived from Arabidopsis thaliana grown under control and abiotic stress conditions, where the organellar transcriptome had not been examined. The most appreciable effects were found for heat stress, which induces a global reduction in splicing and editing efficiency, and leads to increased abundance of chloroplast transcripts, including genic, intergenic, and antisense transcripts. Moreover, by concomitantly analyzing nuclear transcripts that encode chloroplast gene expression regulators from the same libraries, we demonstrate the possibility of achieving a holistic understanding of the nucleus-organelle system. ChloroSeq thus represents a unique method for streamlining RNA-Seq data interpretation of the chloroplast transcriptome and its regulators.
Keywords: Arabidopsis thaliana; RNA editing; RNA splicing; RNA-Seq; chloroplast; heat stress; introns; noncoding RNAs; transcriptome.
Copyright © 2016 Castandet et al.
Figures

References
-
- Allorent G., Courtois F., Chevalier F., Lerbs-Mache S., 2013. Plastid gene expression during chloroplast differentiation and dedifferentiation into non-photosynthetic plastids during seed formation. Plant Mol. Biol. 82(1–2): 59–70. - PubMed
-
- Cahoon A. B., Takacs E. M., Sharpe R. M., Stern D. B., 2008. Nuclear, chloroplast, and mitochondrial transcript abundance along a maize leaf developmental gradient. Plant Mol. Biol. 66(1–2): 33–46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources