

Stimulation of the hypothalamic arcuate nucleus increases brown adipose tissue nerve activity via hypothalamic paraventricular and dorsomedial nuclei
- PMID: 27402666
- PMCID: PMC5008654
- DOI: 10.1152/ajpheart.00176.2016
Stimulation of the hypothalamic arcuate nucleus increases brown adipose tissue nerve activity via hypothalamic paraventricular and dorsomedial nuclei
Abstract
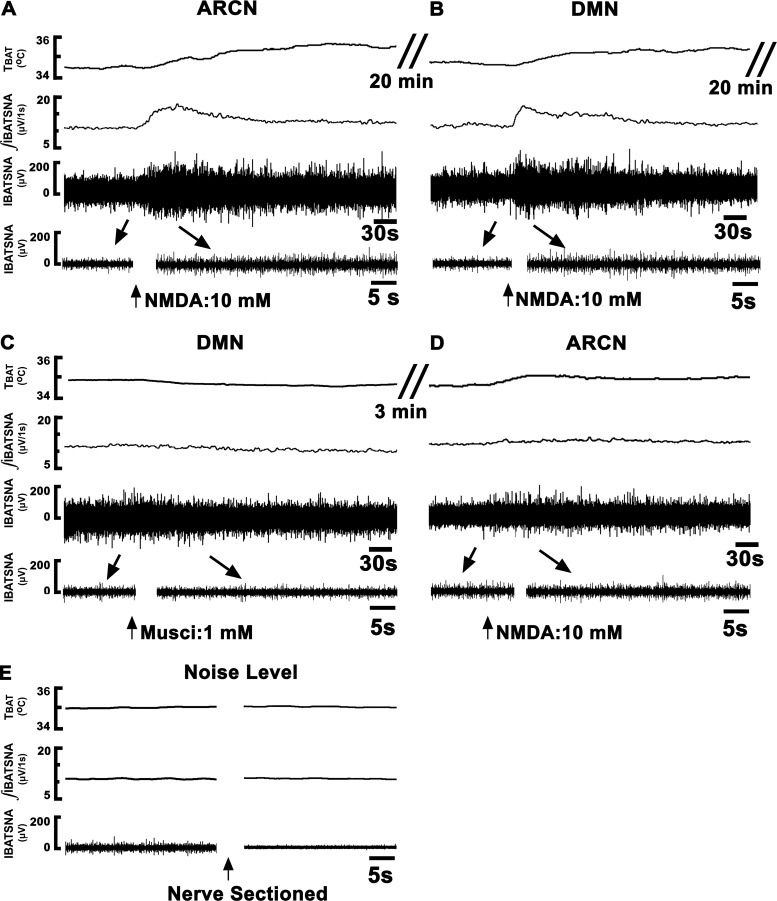
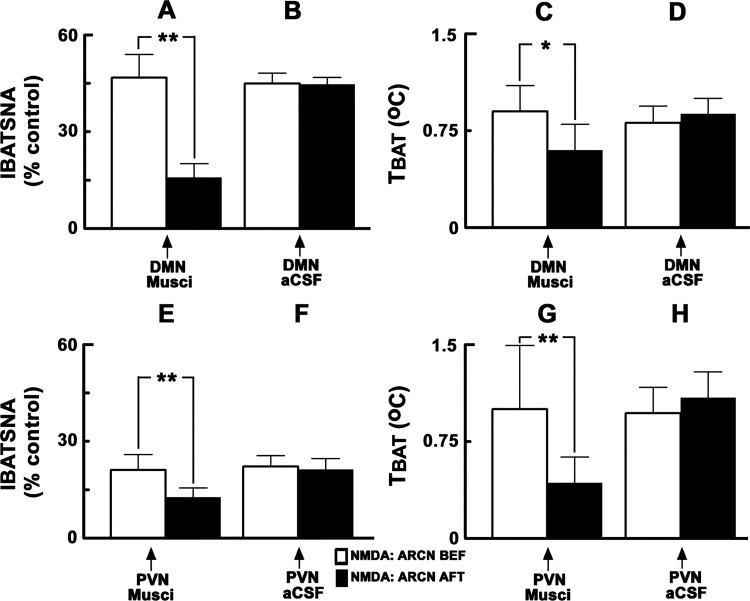
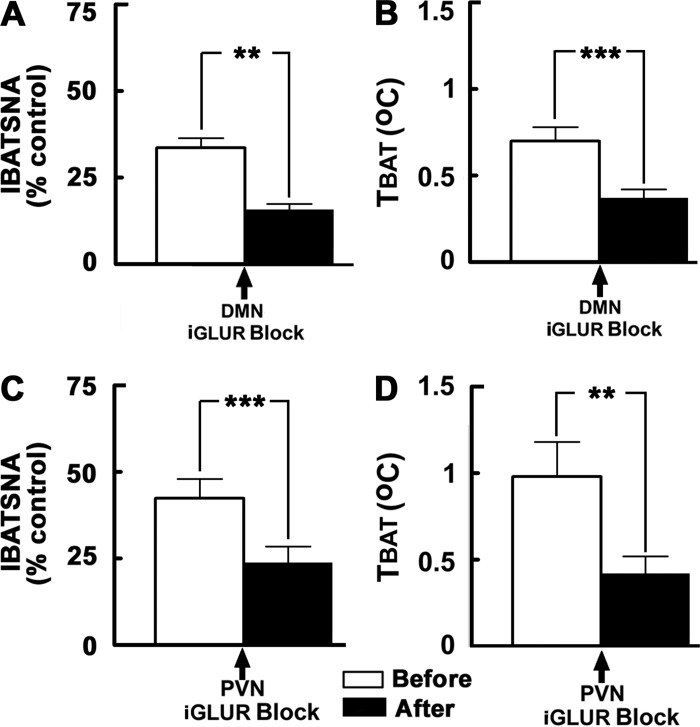
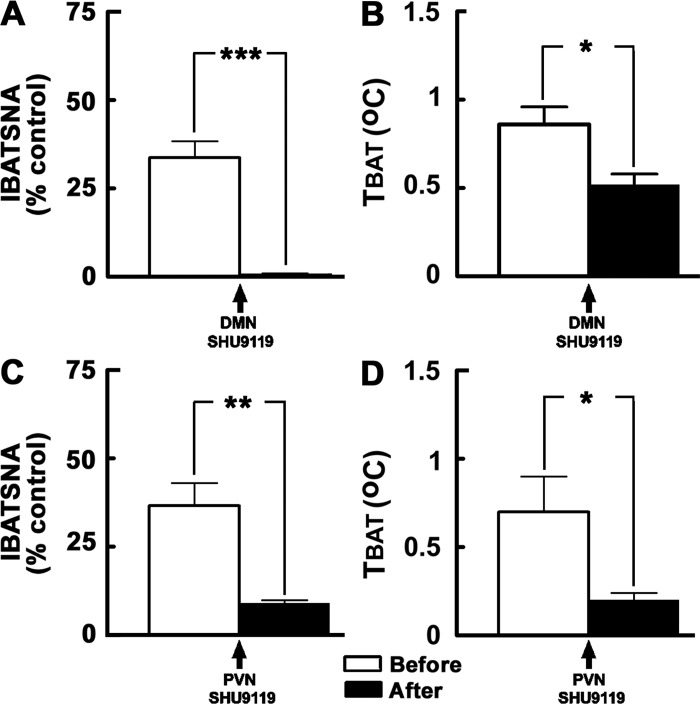
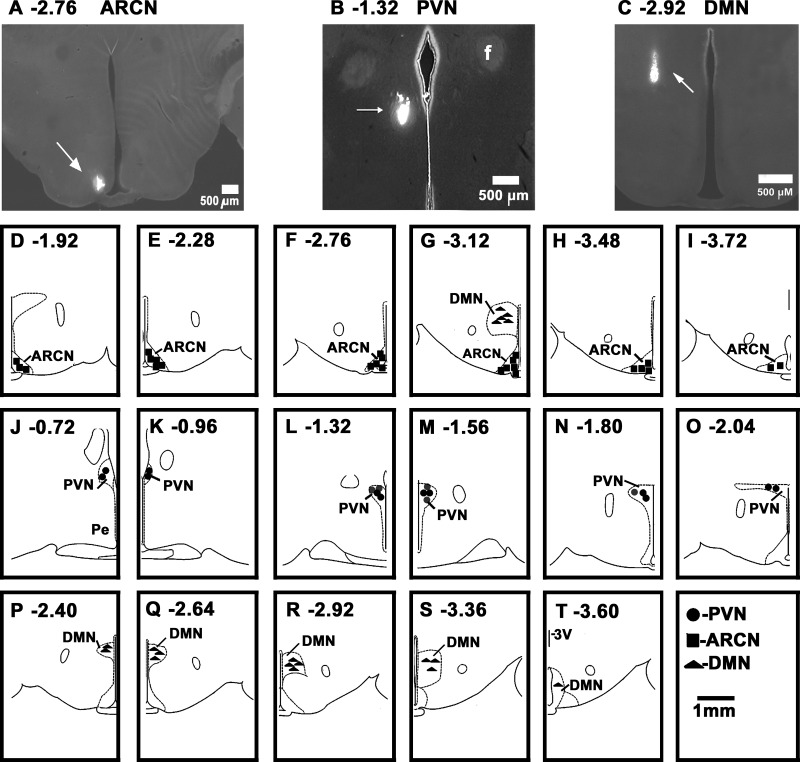
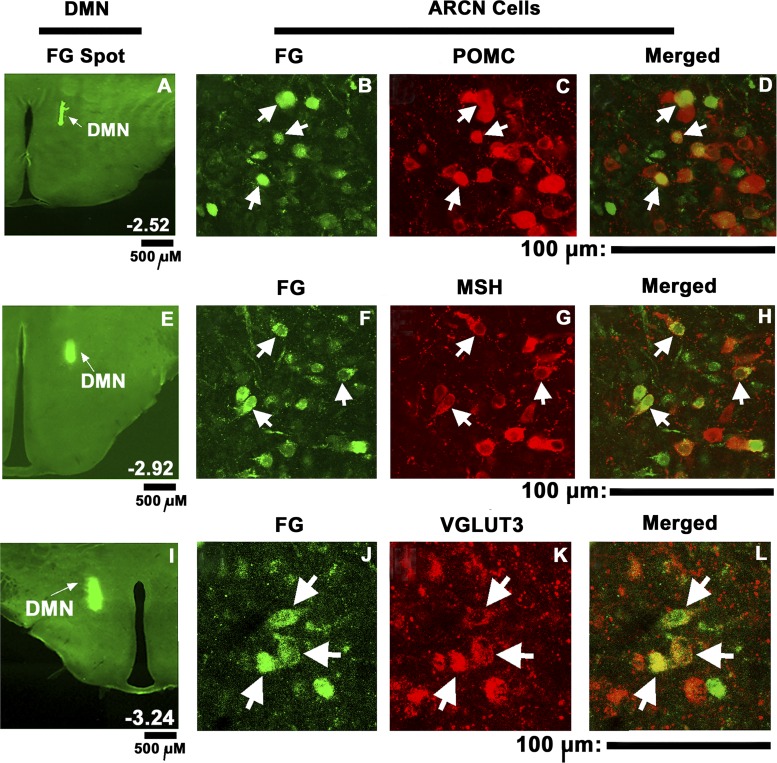
Hypothalamic arcuate nucleus (ARCN) stimulation elicited increases in sympathetic nerve activity (IBATSNA) and temperature (TBAT) of interscapular brown adipose tissue (IBAT). The role of hypothalamic dorsomedial (DMN) and paraventricular (PVN) nuclei in mediating these responses was studied in urethane-anesthetized, artificially ventilated, male Wistar rats. In different groups of rats, inhibition of neurons in the DMN and PVN by microinjections of muscimol attenuated the increases in IBATSNA and TBAT elicited by microinjections of N-methyl-d-aspartic acid into the ipsilateral ARCN. In other groups of rats, blockade of ionotropic glutamate receptors by combined microinjections of D(-)-2-amino-7-phosphono-heptanoic acid (D-AP7) and NBQX into the DMN and PVN attenuated increases in IBATSNA and TBAT elicited by ARCN stimulation. Blockade of melanocortin 3/4 receptors in the DMN and PVN in other groups of rats resulted in attenuation of increases in IBATSNA and TBAT elicited by ipsilateral ARCN stimulation. Microinjections of Fluoro-Gold into the DMN resulted in retrograde labeling of cells in the ipsilateral ARCN, and some of these cells contained proopiomelanocortin (POMC), α-melanocyte-stimulating hormone (α-MSH), or vesicular glutamate transporter-3. Since similar projections from ARCN to the PVN have been reported by us and others, these results indicate that neurons containing POMC, α-MSH, and glutamate project from the ARCN to the DMN and PVN. Stimulation of ARCN results in the release of α-MSH and glutamate in the DMN and PVN which, in turn, cause increases in IBATSNA and TBAT.
Keywords: bat nerve activity; brown adipose tissue temperature; dorsomedial nucleus; glutamate receptors; interscapular brown adipose tissue; melanocortin receptors; paraventricular nucleus; proopiomelanocortin.
Copyright © 2016 the American Physiological Society.
Figures
References
-
- Amir S. Stimulation of the paraventricular nucleus with glutamate activates interscapular brown adipose tissue thermogenesis in rats. Brain Res 508: 152–155, 1990. - PubMed
-
- Bai FL, Yamano M, Shiotani Y, Emson PC, Smith AD, Powell JF, Tohyama M. An arcuato-paraventricular and -dorsomedial hypothalamic neuropeptide Y-containing system which lacks noradrenaline in the rat. Brain Res 331: 172–175, 1985. - PubMed
-
- Baker RA, Herkenham M. Arcuate nucleus neurons that project to the hypothalamic paraventricular nucleus: neuropeptidergic identity and consequences of adrenalectomy on mRNA levels in the rat. J Comp Neurol 358: 518–530, 1995. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous