

Shh and ZRS enhancer colocalisation is specific to the zone of polarising activity
- PMID: 27402708
- PMCID: PMC5004883
- DOI: 10.1242/dev.139188
Shh and ZRS enhancer colocalisation is specific to the zone of polarising activity
Abstract
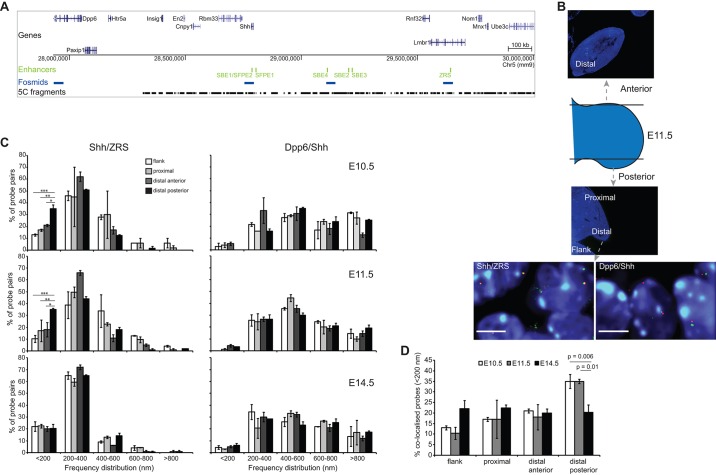
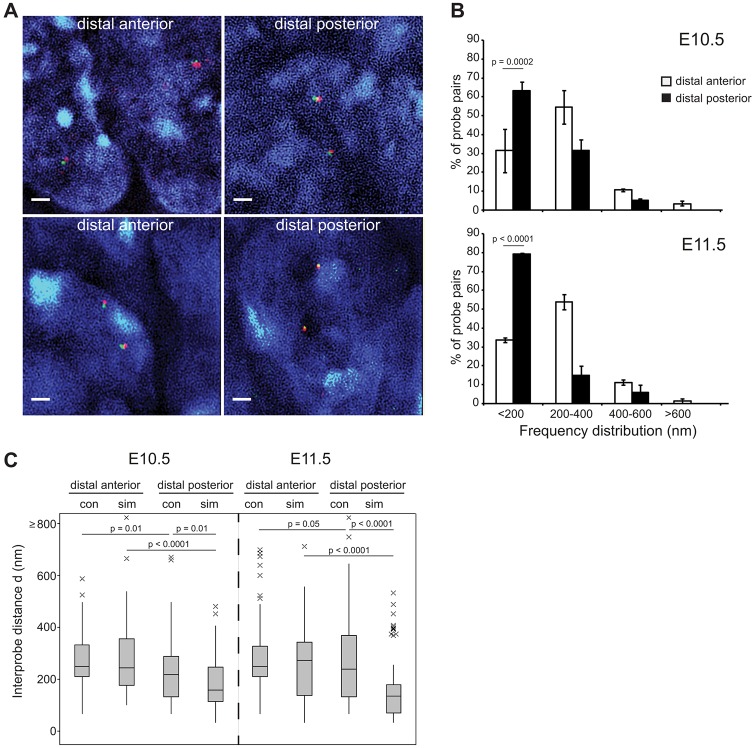
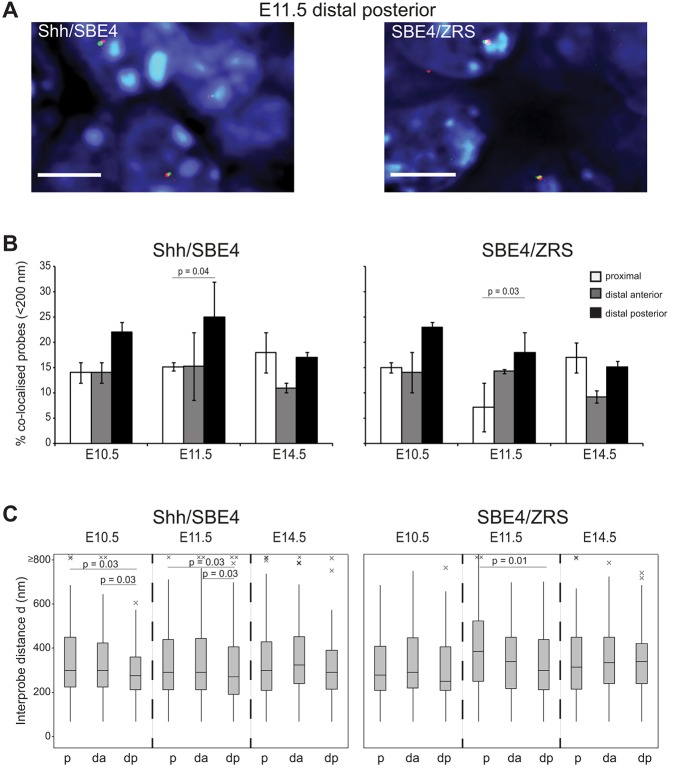
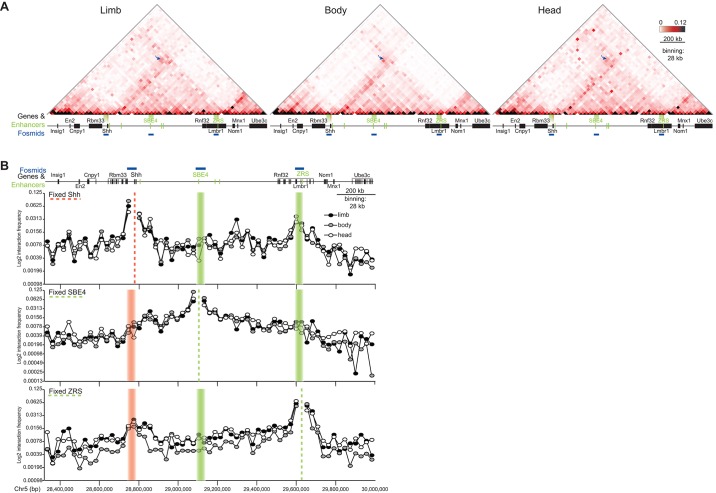
Limb-specific Shh expression is regulated by the (∼1 Mb distant) ZRS enhancer. In the mouse, limb bud-restricted spatiotemporal Shh expression occurs from ∼E10 to E11.5 at the distal posterior margin and is essential for correct autopod formation. Here, we have analysed the higher-order chromatin conformation of Shh in expressing and non-expressing tissues, both by fluorescence in situ hybridisation (FISH) and by chromosome conformation capture (5C). Conventional and super-resolution light microscopy identified significantly elevated frequencies of Shh/ZRS colocalisation only in the Shh-expressing regions of the limb bud, in a conformation consistent with enhancer-promoter loop formation. However, in all tissues and at all developmental stages analysed, Shh-ZRS spatial distances were still consistently shorter than those to a neural enhancer located between Shh and ZRS in the genome. 5C identified a topologically associating domain (TAD) over the Shh/ZRS genomic region and enriched interactions between Shh and ZRS throughout E11.5 embryos. Shh/ZRS colocalisation, therefore, correlates with the spatiotemporal domain of limb bud-specific Shh expression, but close Shh and ZRS proximity in the nucleus occurs regardless of whether the gene or enhancer is active. We suggest that this constrained chromatin configuration optimises the opportunity for the active enhancer to locate and instigate the expression of Shh.
Keywords: 5C; Chromosome loop; Enhancer; Limb development; Super-resolution microscopy.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
