

Pathological ribonuclease H1 causes R-loop depletion and aberrant DNA segregation in mitochondria
- PMID: 27402764
- PMCID: PMC4968715
- DOI: 10.1073/pnas.1600537113
Pathological ribonuclease H1 causes R-loop depletion and aberrant DNA segregation in mitochondria
Abstract
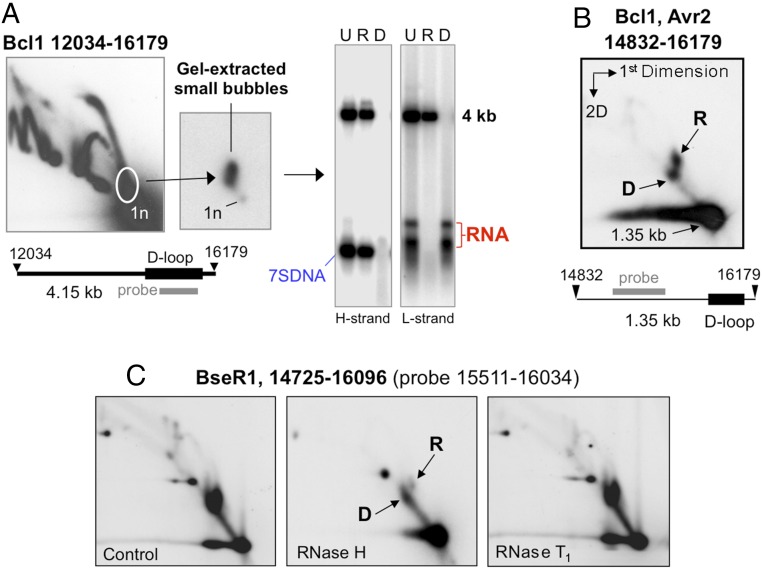
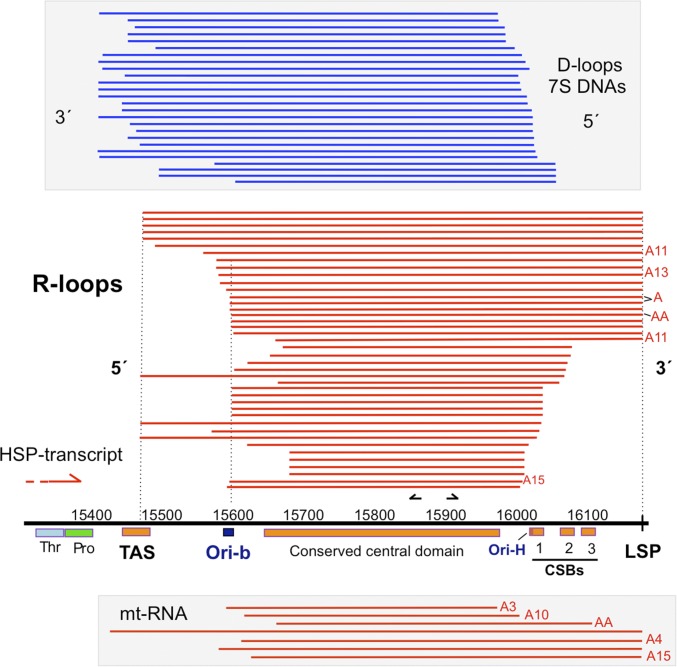
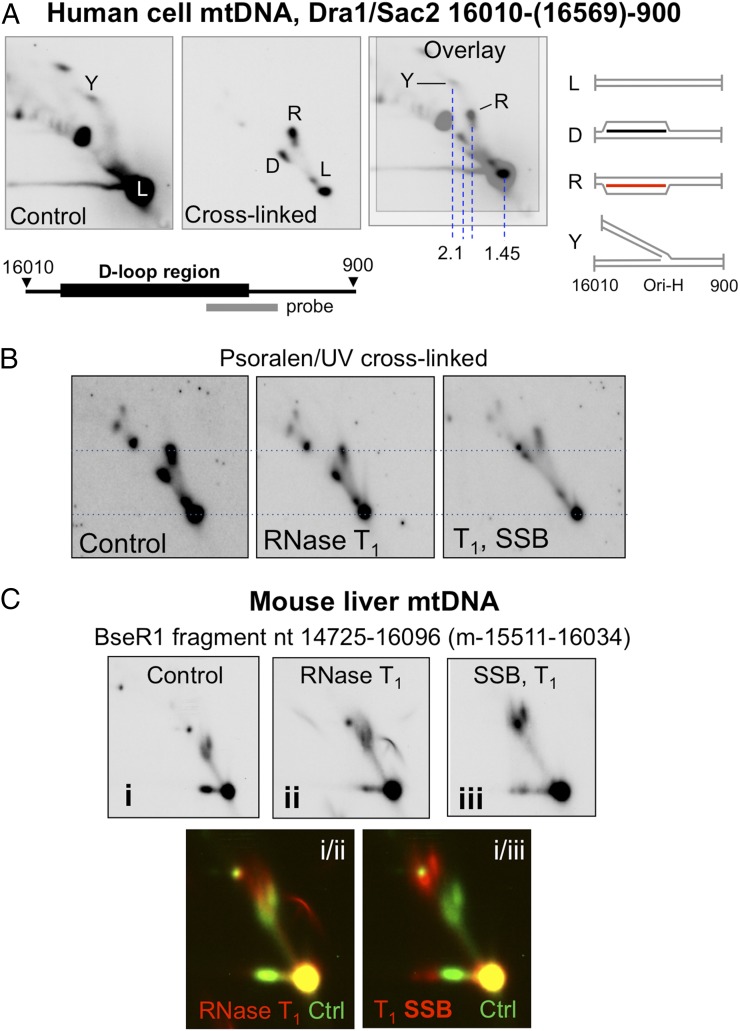
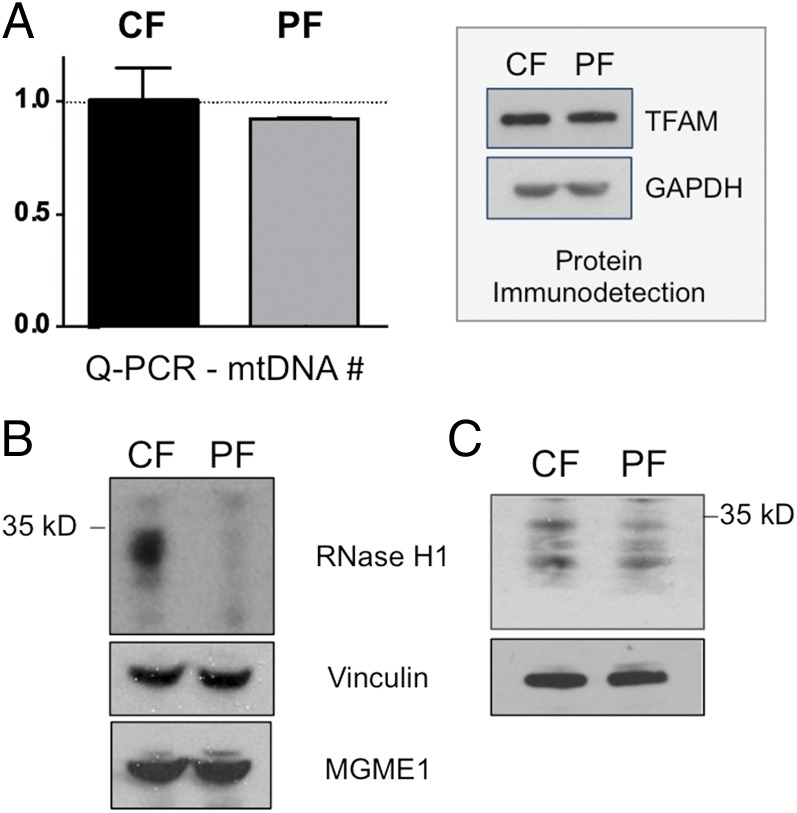
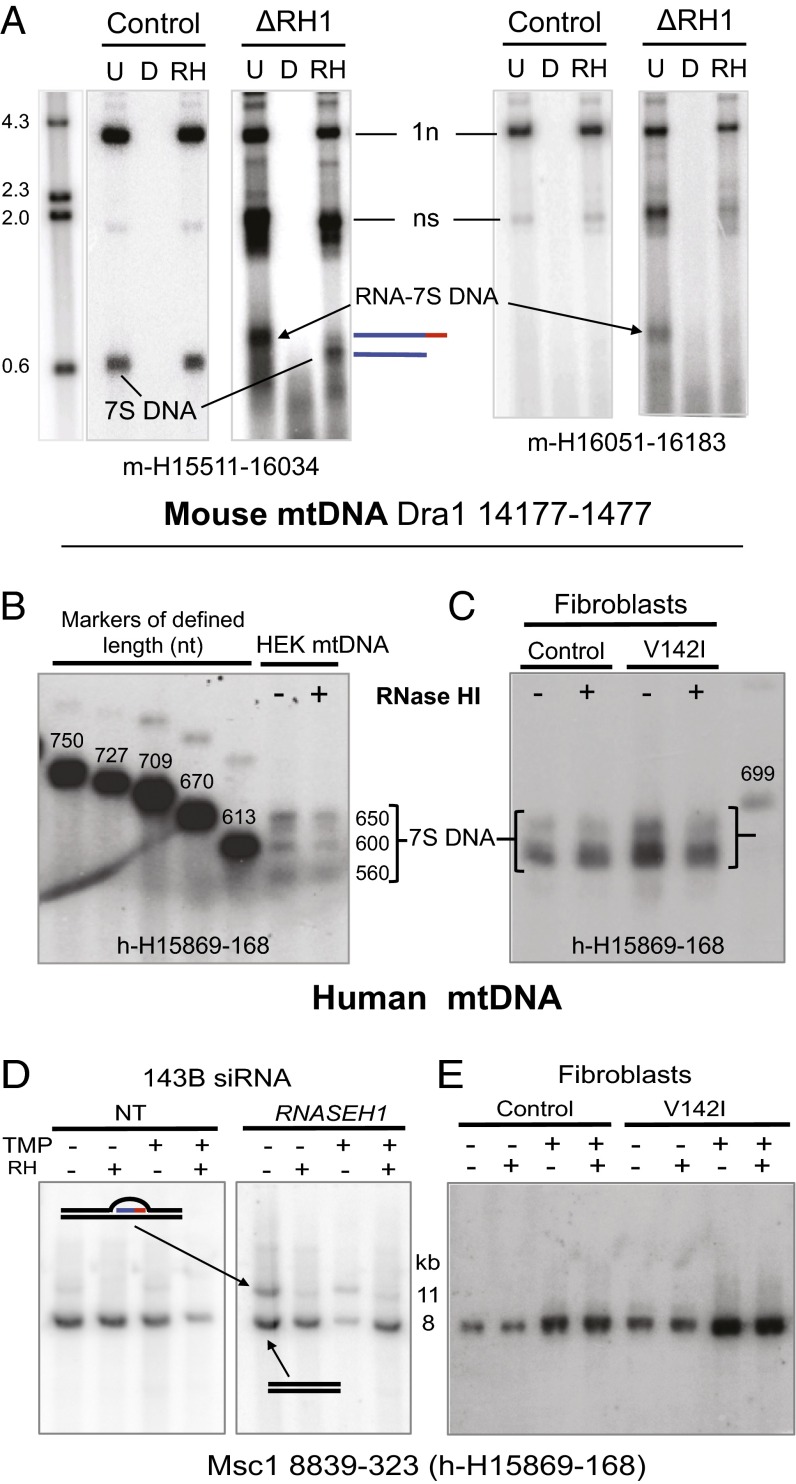
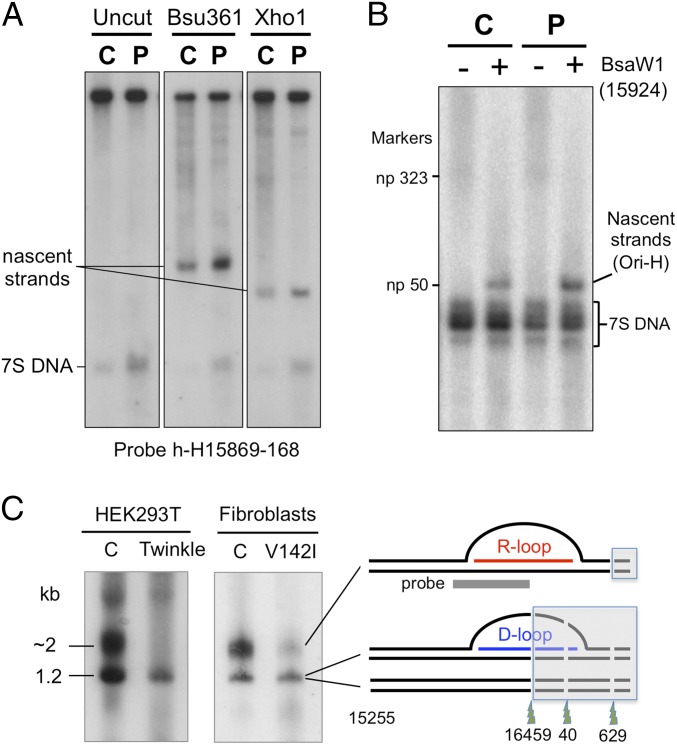
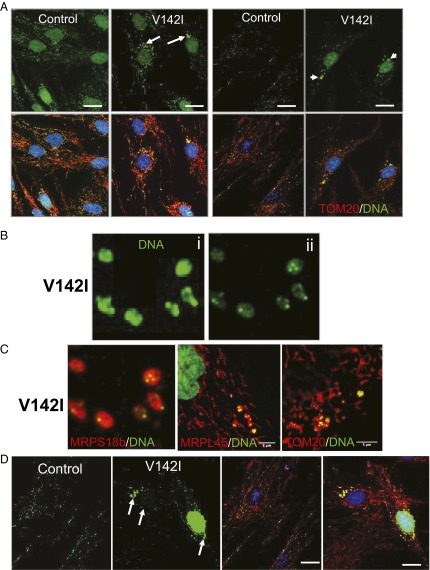
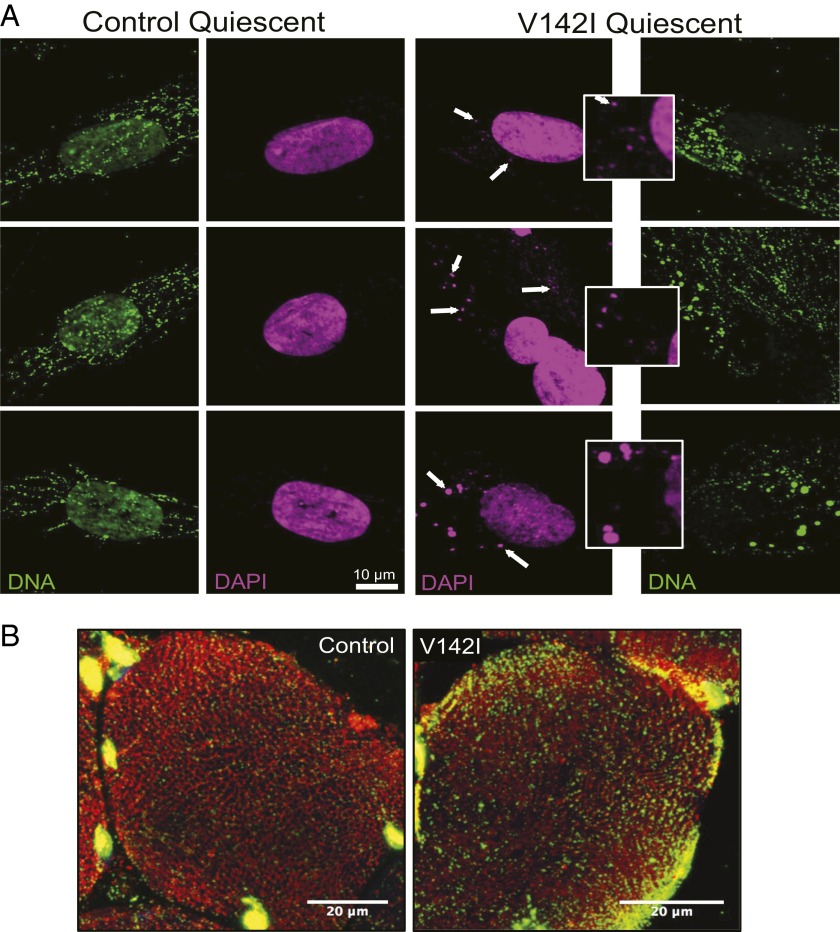
The genetic information in mammalian mitochondrial DNA is densely packed; there are no introns and only one sizeable noncoding, or control, region containing key cis-elements for its replication and expression. Many molecules of mitochondrial DNA bear a third strand of DNA, known as "7S DNA," which forms a displacement (D-) loop in the control region. Here we show that many other molecules contain RNA as a third strand. The RNA of these R-loops maps to the control region of the mitochondrial DNA and is complementary to 7S DNA. Ribonuclease H1 is essential for mitochondrial DNA replication; it degrades RNA hybridized to DNA, so the R-loop is a potential substrate. In cells with a pathological variant of ribonuclease H1 associated with mitochondrial disease, R-loops are of low abundance, and there is mitochondrial DNA aggregation. These findings implicate ribonuclease H1 and RNA in the physical segregation of mitochondrial DNA, perturbation of which represents a previously unidentified disease mechanism.
Keywords: DNA segregation; R-loop; RNase H1; mitochondrial DNA; mitochondrial disease.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
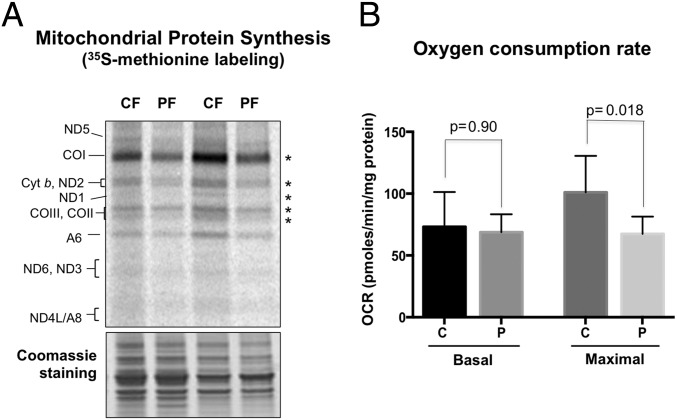
References
-
- Area-Gomez E, Schon EA. Mitochondrial genetics and disease. J Child Neurol. 2014;29(9):1208–1215. - PubMed
-
- Kasamatsu H, Vinograd J. Unidirectionality of replication in mouse mitochondrial DNA. Nat New Biol. 1973;241(108):103–105. - PubMed
-
- Yasukawa T, Yang MY, Jacobs HT, Holt IJ. A bidirectional origin of replication maps to the major noncoding region of human mitochondrial DNA. Mol Cell. 2005;18(6):651–662. - PubMed
-
- Fish J, Raule N, Attardi G. Discovery of a major D-loop replication origin reveals two modes of human mtDNA synthesis. Science. 2004;306(5704):2098–2101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
