

Protease-activated Receptor-4 Signaling and Trafficking Is Regulated by the Clathrin Adaptor Protein Complex-2 Independent of β-Arrestins
- PMID: 27402844
- PMCID: PMC5000090
- DOI: 10.1074/jbc.M116.729285
Protease-activated Receptor-4 Signaling and Trafficking Is Regulated by the Clathrin Adaptor Protein Complex-2 Independent of β-Arrestins
Abstract
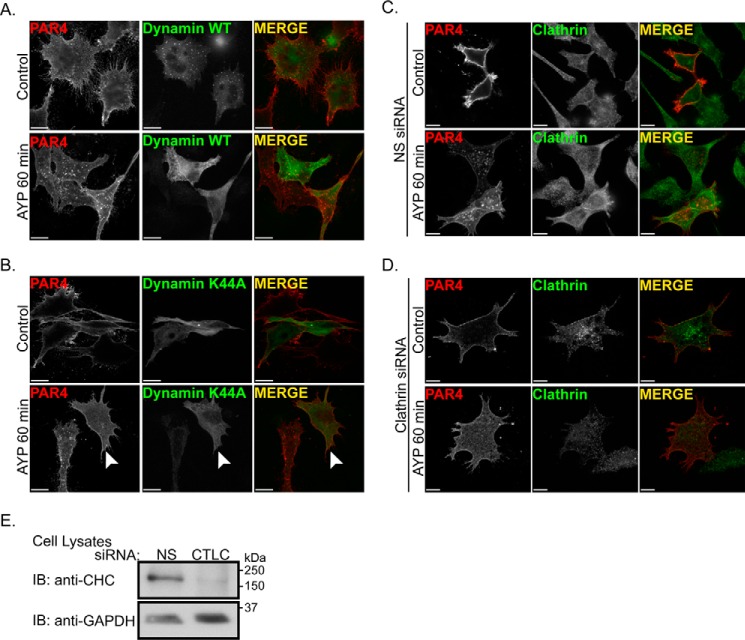
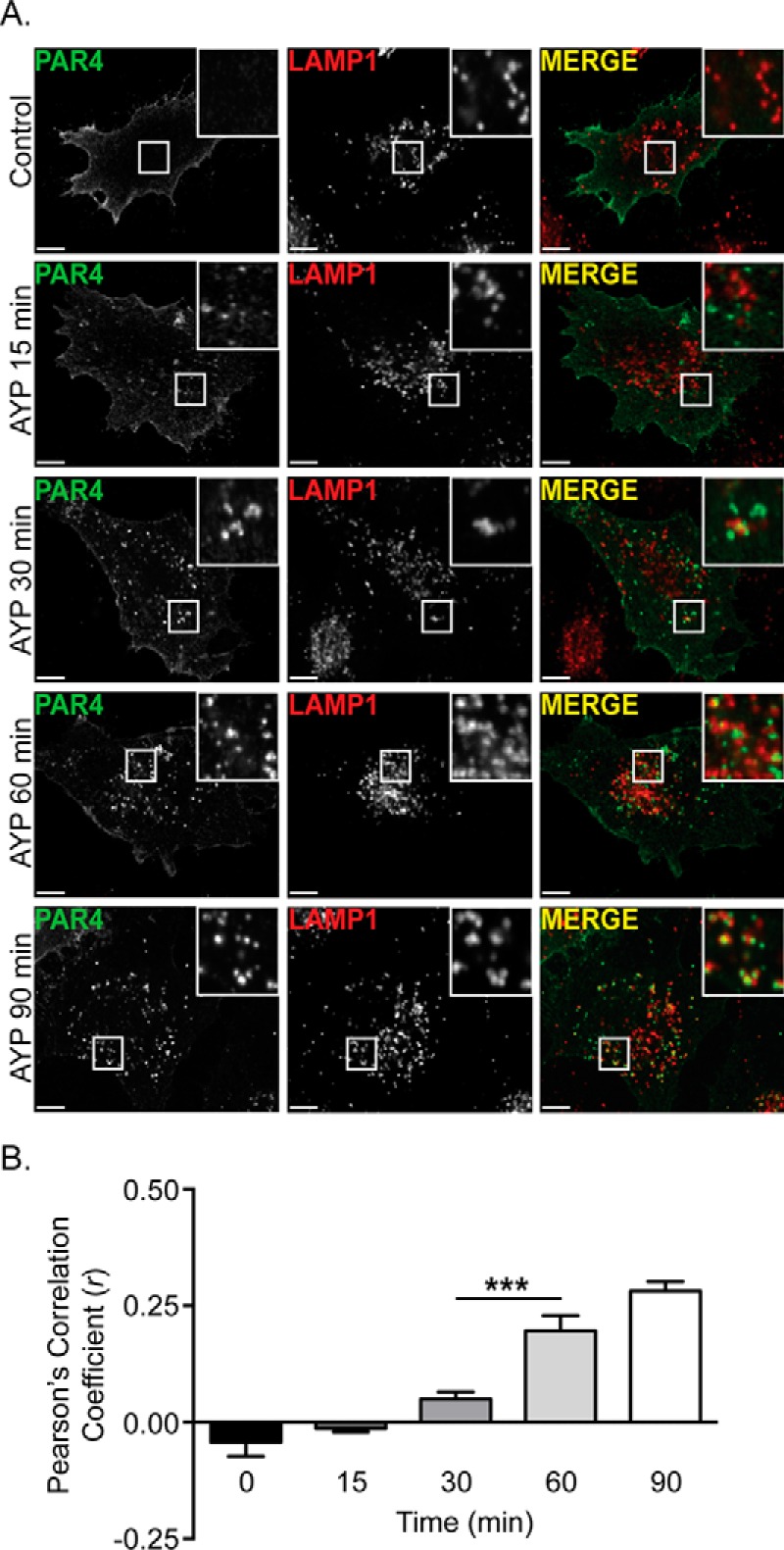
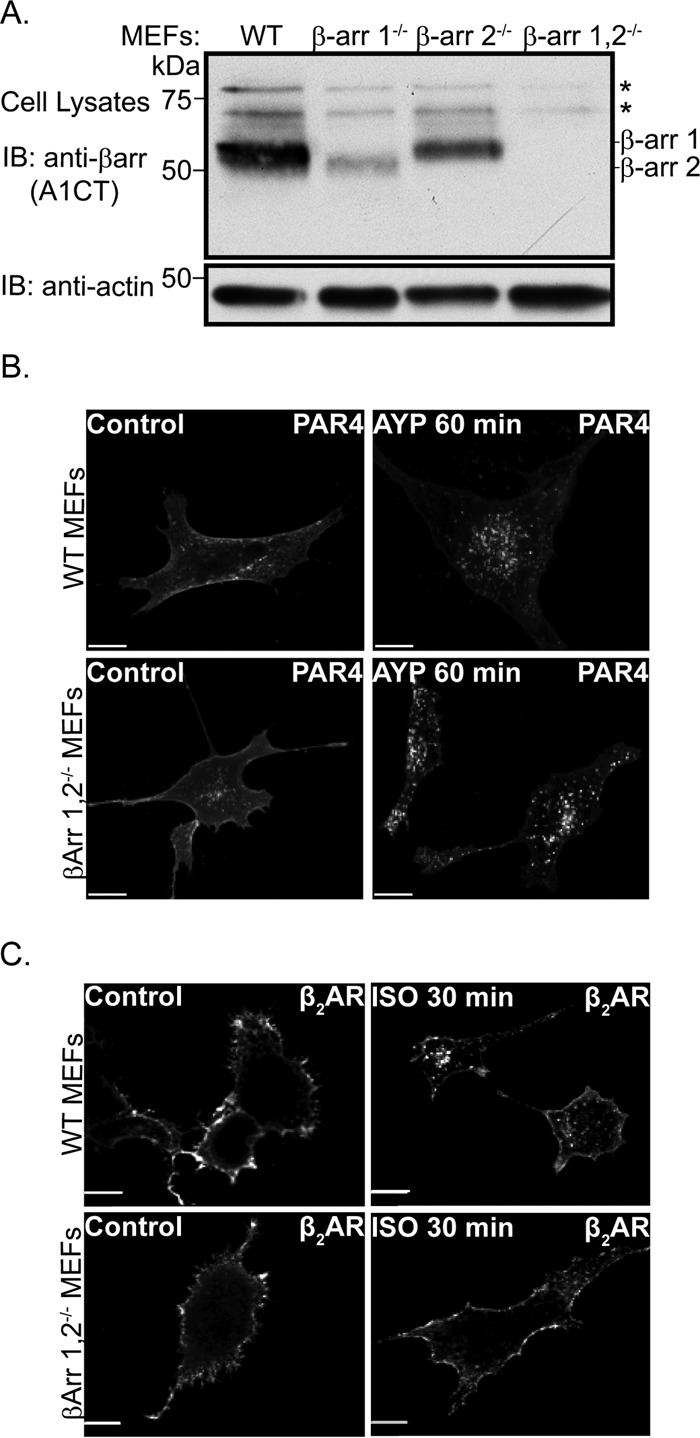
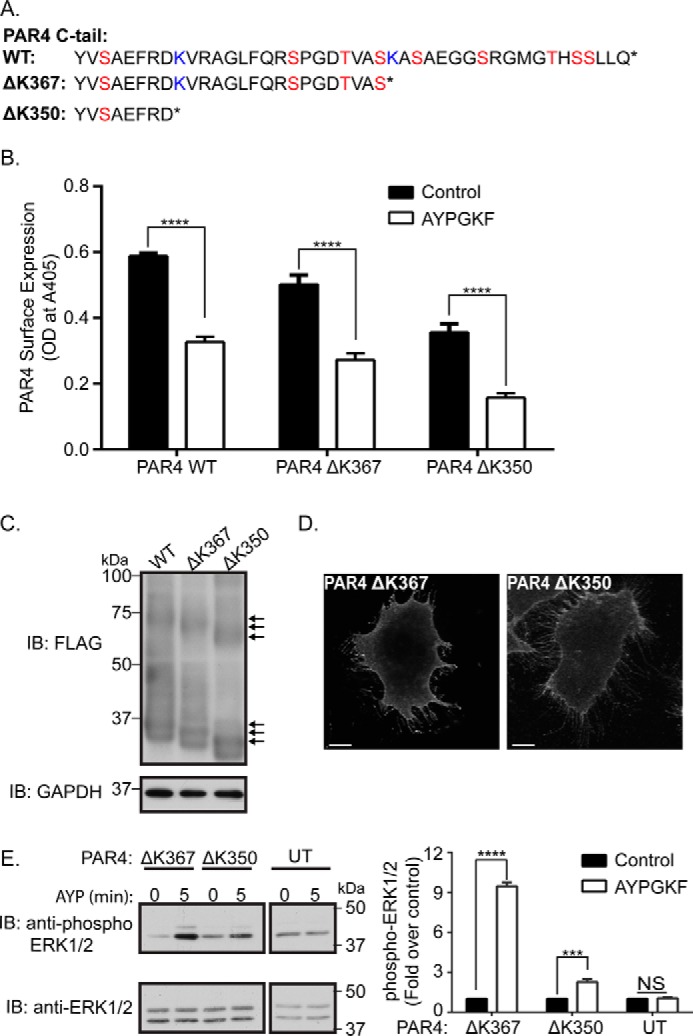
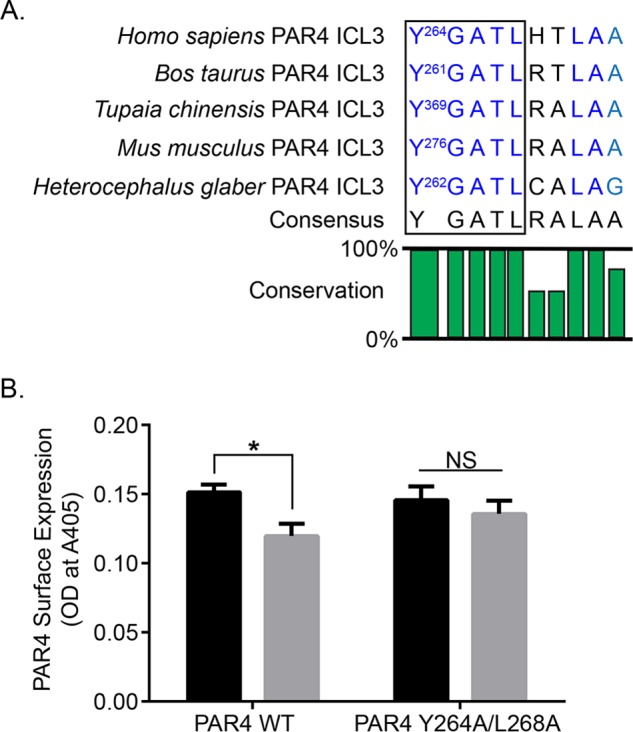
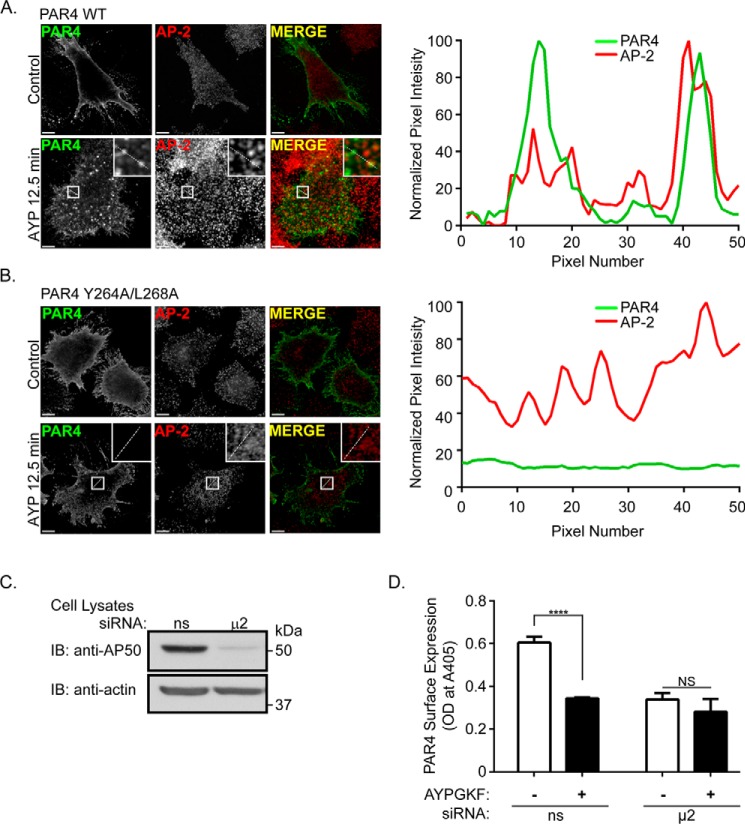
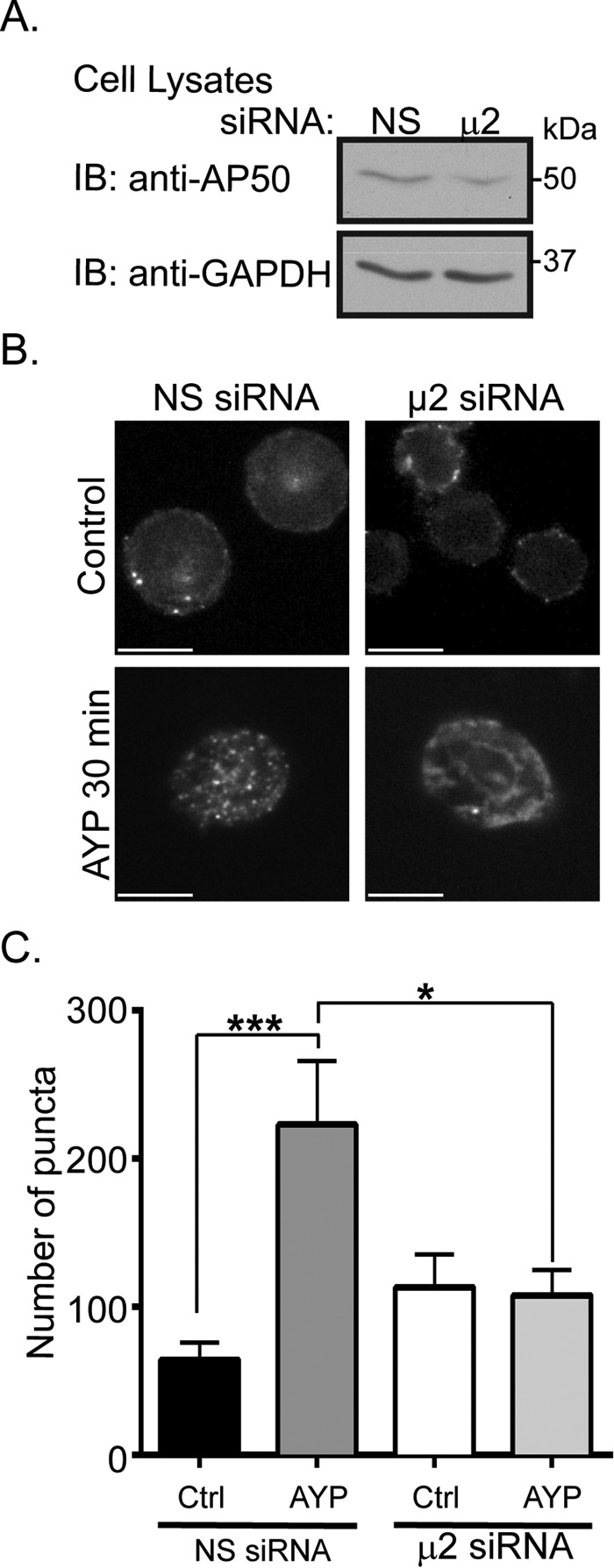
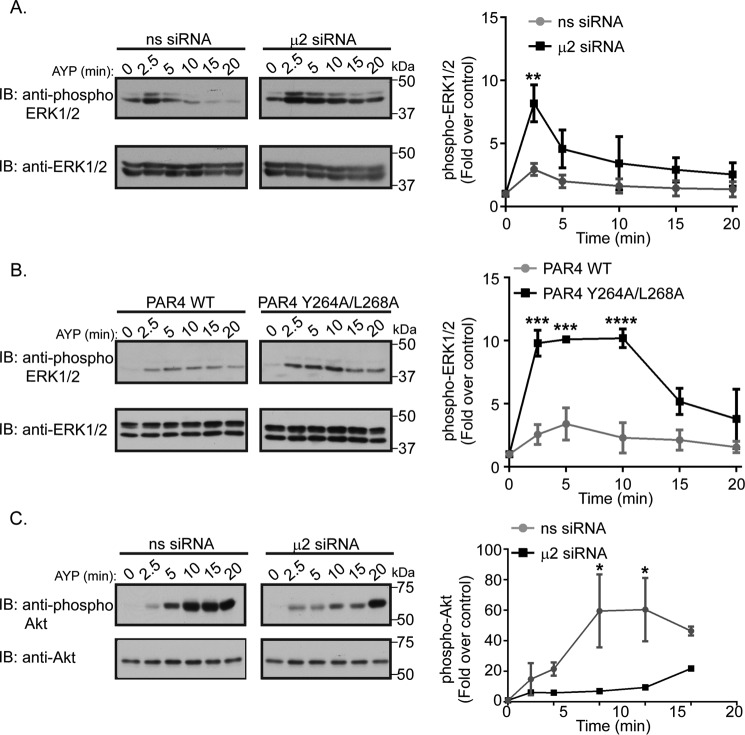
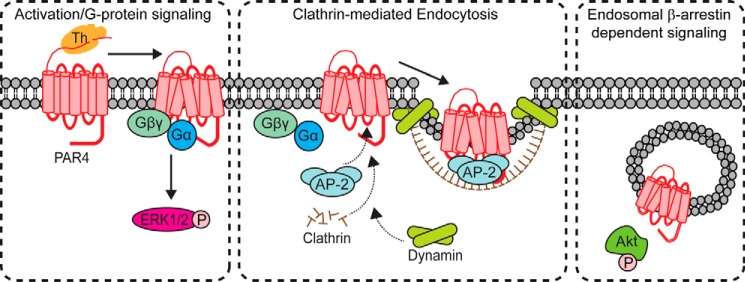
Protease-activated receptor-4 (PAR4) is a G protein-coupled receptor (GPCR) for thrombin and is proteolytically activated, similar to the prototypical PAR1. Due to the irreversible activation of PAR1, receptor trafficking is intimately linked to signal regulation. However, unlike PAR1, the mechanisms that control PAR4 trafficking are not known. Here, we sought to define the mechanisms that control PAR4 trafficking and signaling. In HeLa cells depleted of clathrin by siRNA, activated PAR4 failed to internalize. Consistent with clathrin-mediated endocytosis, expression of a dynamin dominant-negative K44A mutant also blocked activated PAR4 internalization. However, unlike most GPCRs, PAR4 internalization occurred independently of β-arrestins and the receptor's C-tail domain. Rather, we discovered a highly conserved tyrosine-based motif in the third intracellular loop of PAR4 and found that the clathrin adaptor protein complex-2 (AP-2) is important for internalization. Depletion of AP-2 inhibited PAR4 internalization induced by agonist. In addition, mutation of the critical residues of the tyrosine-based motif disrupted agonist-induced PAR4 internalization. Using Dami megakaryocytic cells, we confirmed that AP-2 is required for agonist-induced internalization of endogenous PAR4. Moreover, inhibition of activated PAR4 internalization enhanced ERK1/2 signaling, whereas Akt signaling was markedly diminished. These findings indicate that activated PAR4 internalization requires AP-2 and a tyrosine-based motif and occurs independent of β-arrestins, unlike most classical GPCRs. Moreover, these findings are the first to show that internalization of activated PAR4 is linked to proper ERK1/2 and Akt activation.
Keywords: Akt; G protein-coupled receptor (GPCR); adaptor protein complex-2; arrestin; clathrin; extracellular-signal-regulated kinase (ERK); lysosome; megakaryocyte; thrombin.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Coughlin S. R. (2005) Protease-activated receptors in hemostasis, thrombosis, and vascular biology. J. Thromb. Haemost. 3, 1800–1814 - PubMed
-
- Vu T. K., Hung D. T., Wheaton V. I., and Coughlin S. R. (1991) Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell 64, 1057–1068 - PubMed
-
- Vu T. K., Wheaton V. I., Hung D. T., Charo I., and Coughlin S. R. (1991) Domains specifying thrombin-receptor interaction. Nature 353, 674–677 - PubMed
-
- Scarborough R. M., Naughton M. A., Teng W., Hung D. T., Rose J., Vu T. K., Wheaton V. I., Turck C. W., and Coughlin S. R. (1992) Tethered ligand agonist peptides: structural requirements for thrombin receptor activation reveal mechanism of proteolytic unmasking of agonist function. J. Biol. Chem. 267, 13146–13149 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
