

Electrochemical Evidence for Neuroglobin Activity on NO at Physiological Concentrations
- PMID: 27402851
- PMCID: PMC5009268
- DOI: 10.1074/jbc.M116.730176
Electrochemical Evidence for Neuroglobin Activity on NO at Physiological Concentrations
Abstract
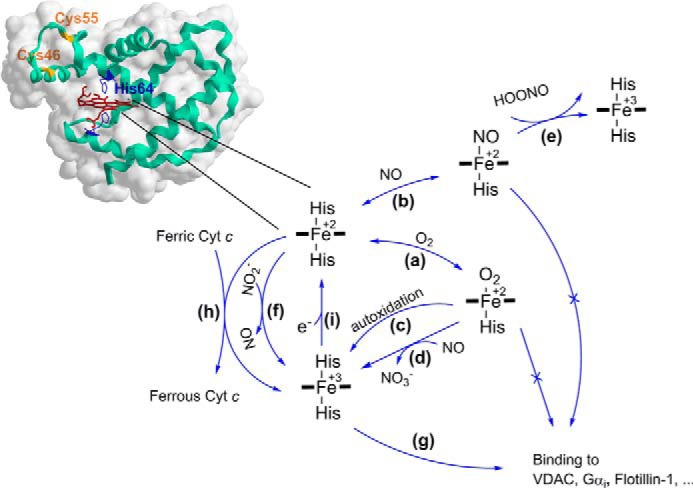
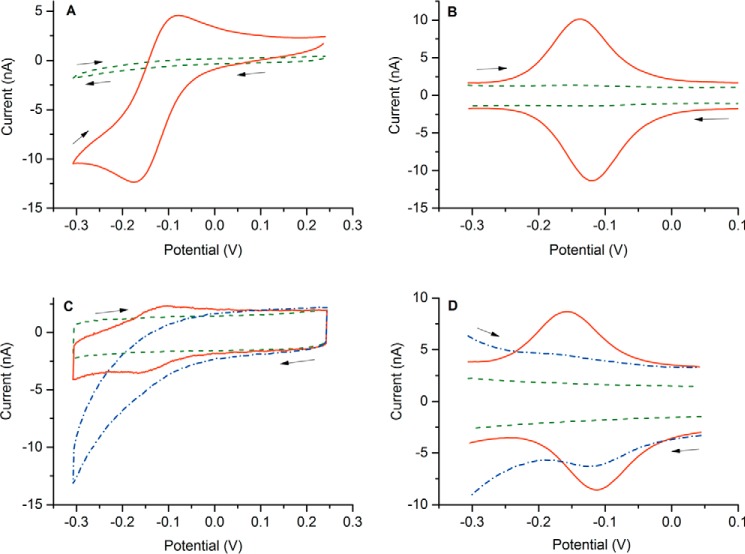
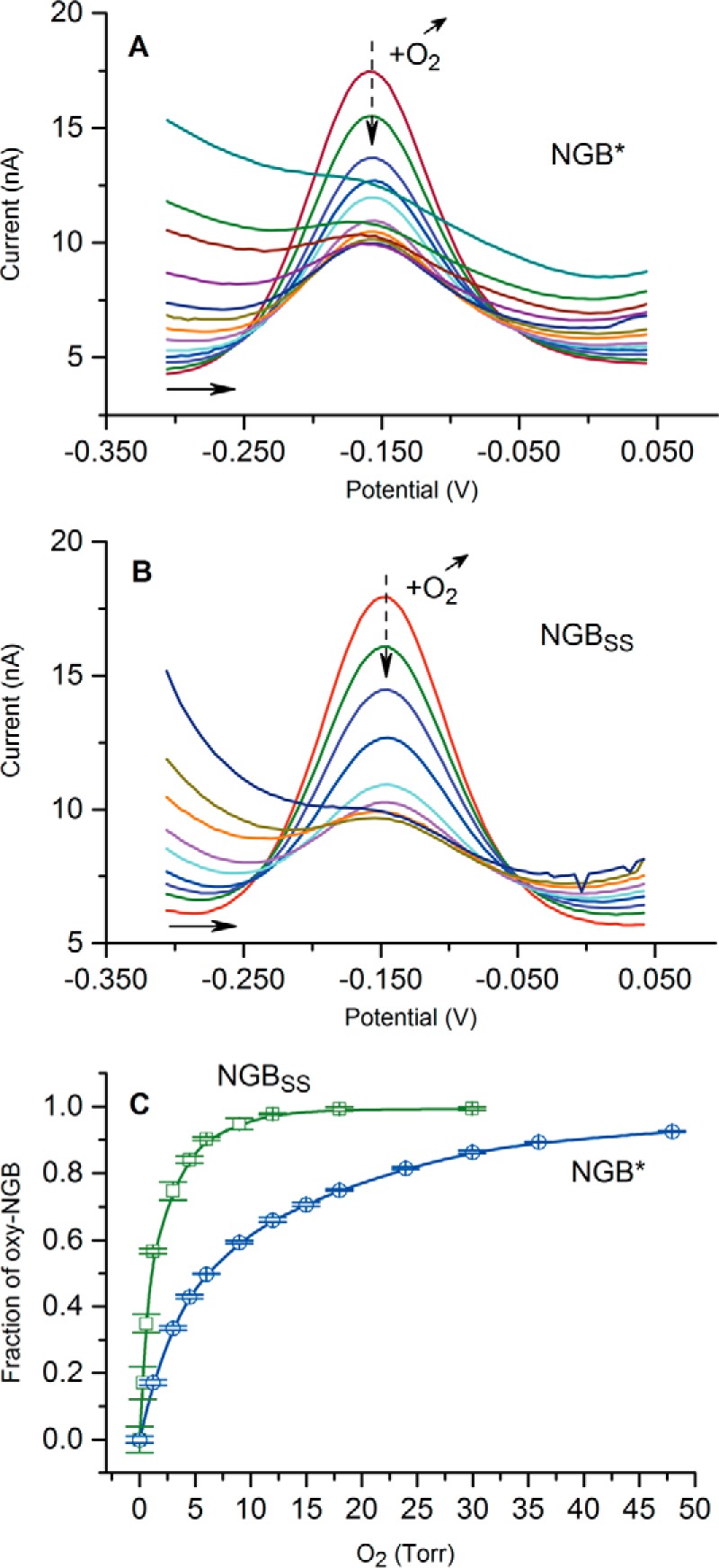
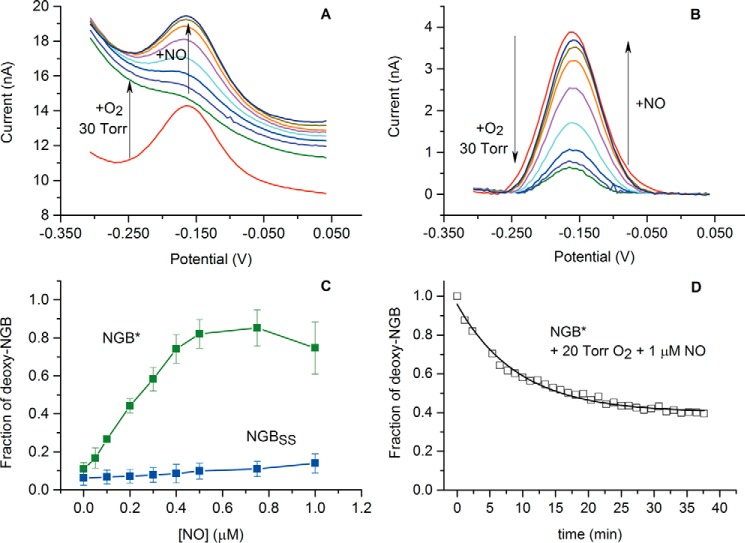
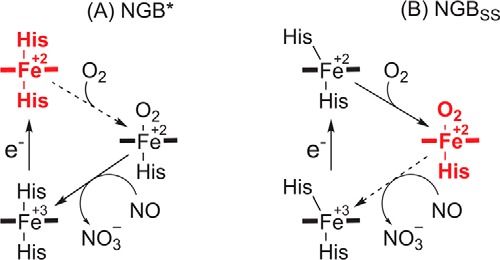
The true function of neuroglobin (Ngb) and, particularly, human Ngb (NGB) has been under debate since its discovery 15 years ago. It has been expected to play a role in oxygen binding/supply, but a variety of other functions have been put forward, including NO dioxygenase activity. However, in vitro studies that could unravel these potential roles have been hampered by the lack of an Ngb-specific reductase. In this work, we used electrochemical measurements to investigate the role of an intermittent internal disulfide bridge in determining NO oxidation kinetics at physiological NO concentrations. The use of a polarized electrode to efficiently interconvert the ferric (Fe(3+)) and ferrous (Fe(2+)) forms of an immobilized NGB showed that the disulfide bridge both defines the kinetics of NO dioxygenase activity and regulates appearance of the free ferrous deoxy-NGB, which is the redox active form of the protein in contrast to oxy-NGB. Our studies further identified a role for the distal histidine, interacting with the hexacoordinated iron atom of the heme, in oxidation kinetics. These findings may be relevant in vivo, for example, in blocking apoptosis by reduction of ferric cytochrome c, and gentle tuning of NO concentration in the tissues.
Keywords: electrochemistry; electron transfer; neuroglobin; nitric oxide; oxygen binding; protein chemistry.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
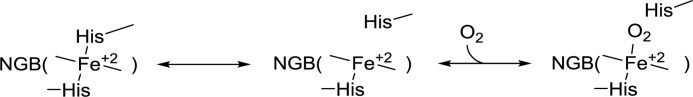
References
-
- Burmester T., Weich B., Reinhardt S., and Hankeln T. (2000) A vertebrate globin expressed in the brain. Nature 407, 520–523 - PubMed
-
- Burmester T., and Hankeln T. (2014) Function and evolution of vertebrate globins. Acta Physiol. 211, 501–514 - PubMed
-
- Ascenzi P., Gustincich S., and Marino M. (2014) Mammalian nerve globins in search of functions. IUBMB Life 66, 268–276 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
