

Effect of BET Missense Mutations on Bromodomain Function, Inhibitor Binding and Stability
- PMID: 27403962
- PMCID: PMC4942050
- DOI: 10.1371/journal.pone.0159180
Effect of BET Missense Mutations on Bromodomain Function, Inhibitor Binding and Stability
Abstract
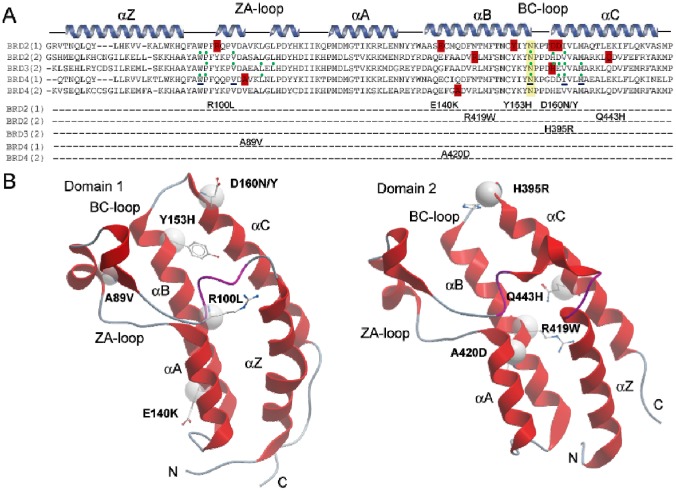
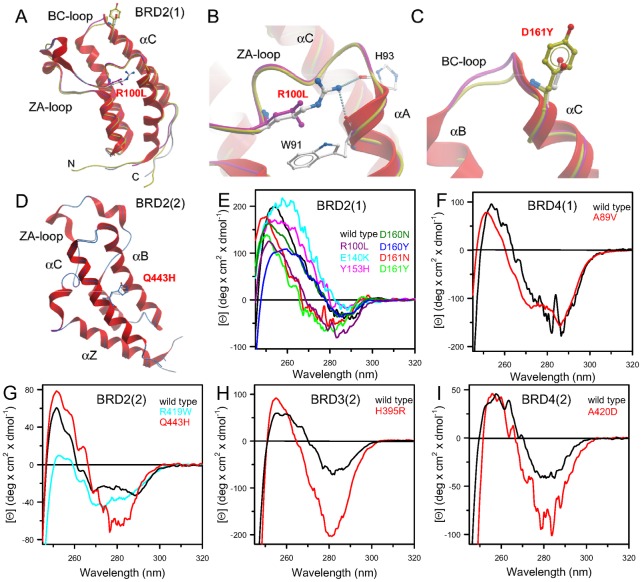
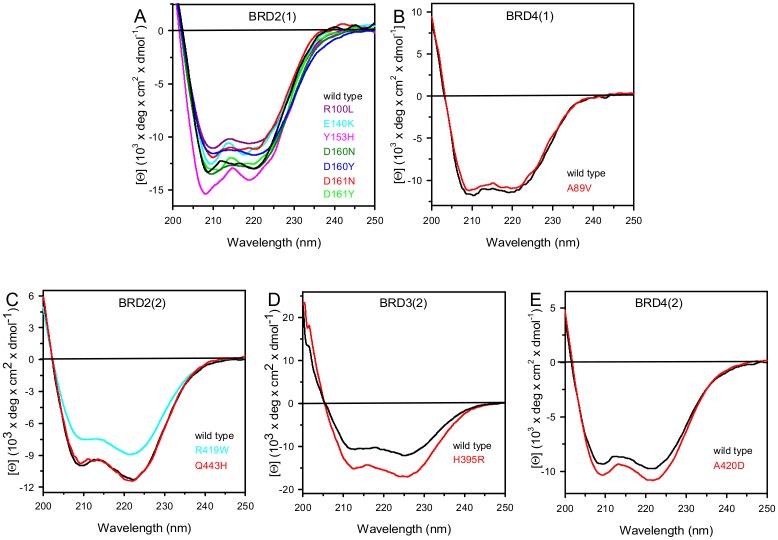
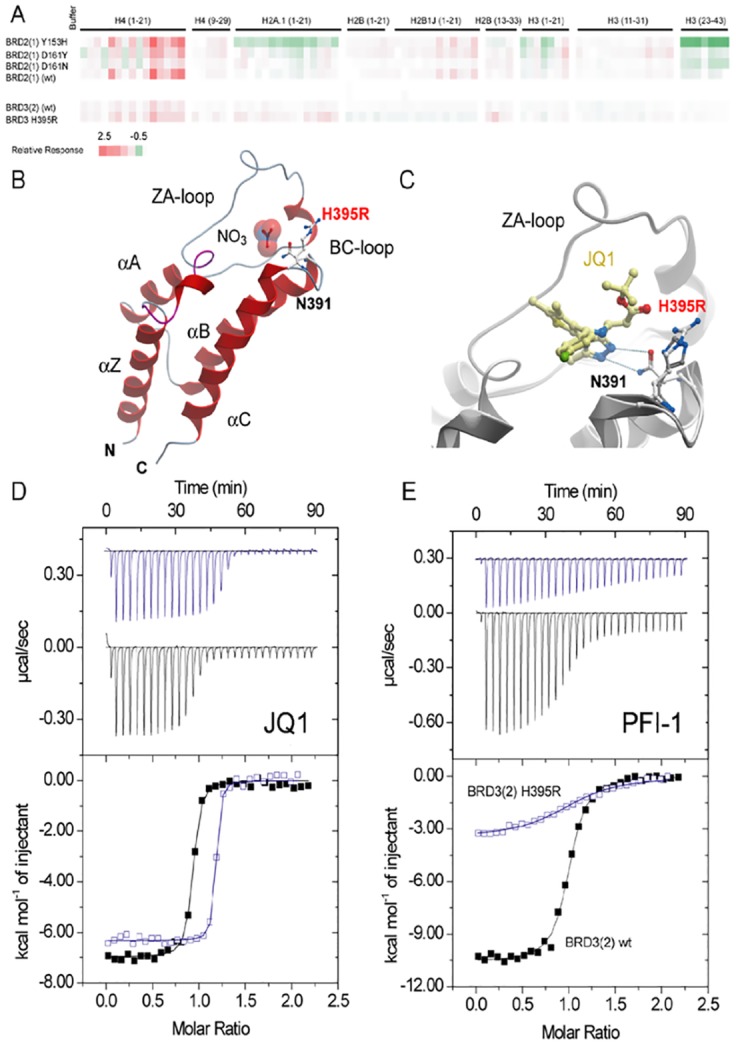
Lysine acetylation is an important epigenetic mark regulating gene transcription and chromatin structure. Acetylated lysine residues are specifically recognized by bromodomains, small protein interaction modules that read these modification in a sequence and acetylation dependent way regulating the recruitment of transcriptional regulators and chromatin remodelling enzymes to acetylated sites in chromatin. Recent studies revealed that bromodomains are highly druggable protein interaction domains resulting in the development of a large number of bromodomain inhibitors. BET bromodomain inhibitors received a lot of attention in the oncology field resulting in the rapid translation of early BET bromodomain inhibitors into clinical studies. Here we investigated the effects of mutations present as polymorphism or found in cancer on BET bromodomain function and stability and the influence of these mutants on inhibitor binding. We found that most BET missense mutations localize to peripheral residues in the two terminal helices. Crystal structures showed that the three dimensional structure is not compromised by these mutations but mutations located in close proximity to the acetyl-lysine binding site modulate acetyl-lysine and inhibitor binding. Most mutations affect significantly protein stability and tertiary structure in solution, suggesting new interactions and an alternative network of protein-protein interconnection as a consequence of single amino acid substitution. To our knowledge this is the first report studying the effect of mutations on bromodomain function and inhibitor binding.
Conflict of interest statement
Figures
Similar articles
-
Small molecule inhibitors of bromodomain-acetyl-lysine interactions.ACS Chem Biol. 2015 Jan 16;10(1):22-39. doi: 10.1021/cb500996u. ACS Chem Biol. 2015. PMID: 25549280 Review.
-
The Bromodomain and Extra-Terminal Domain (BET) Family: Functional Anatomy of BET Paralogous Proteins.Int J Mol Sci. 2016 Nov 7;17(11):1849. doi: 10.3390/ijms17111849. Int J Mol Sci. 2016. PMID: 27827996 Free PMC article. Review.
-
Covalent-Fragment Screening of BRD4 Identifies a Ligandable Site Orthogonal to the Acetyl-Lysine Binding Sites.ACS Chem Biol. 2020 Apr 17;15(4):1036-1049. doi: 10.1021/acschembio.0c00058. Epub 2020 Mar 23. ACS Chem Biol. 2020. PMID: 32149490 Free PMC article.
-
Structural ramification for acetyl-lysine recognition by the bromodomain of human BRG1 protein, a central ATPase of the SWI/SNF remodeling complex.Chembiochem. 2007 Jul 23;8(11):1308-16. doi: 10.1002/cbic.200600562. Chembiochem. 2007. PMID: 17582821
-
Cancer-associated polybromo-1 bromodomain 4 missense variants variably impact bromodomain ligand binding and cell growth suppression.J Biol Chem. 2024 Apr;300(4):107146. doi: 10.1016/j.jbc.2024.107146. Epub 2024 Mar 7. J Biol Chem. 2024. PMID: 38460939 Free PMC article.
Cited by
-
Bromodomain Interactions with Acetylated Histone 4 Peptides in the BRD4 Tandem Domain: Effects on Domain Dynamics and Internal Flexibility.Biochemistry. 2022 Nov 1;61(21):2303-2318. doi: 10.1021/acs.biochem.2c00226. Epub 2022 Oct 10. Biochemistry. 2022. PMID: 36215732 Free PMC article.
-
Characterization of human frataxin missense variants in cancer tissues.Hum Mutat. 2019 Sep;40(9):1400-1413. doi: 10.1002/humu.23789. Epub 2019 Jun 18. Hum Mutat. 2019. PMID: 31074541 Free PMC article.
-
Discovery of a hidden transient state in all bromodomain families.Proc Natl Acad Sci U S A. 2021 Jan 26;118(4):e2017427118. doi: 10.1073/pnas.2017427118. Proc Natl Acad Sci U S A. 2021. PMID: 33468647 Free PMC article.
-
Emerging roles of and therapeutic strategies targeting BRD4 in cancer.Cell Immunol. 2019 Mar;337:48-53. doi: 10.1016/j.cellimm.2019.02.001. Epub 2019 Feb 4. Cell Immunol. 2019. PMID: 30832981 Free PMC article. Review.
-
The phosphoglycerate kinase 1 variants found in carcinoma cells display different catalytic activity and conformational stability compared to the native enzyme.PLoS One. 2018 Jul 11;13(7):e0199191. doi: 10.1371/journal.pone.0199191. eCollection 2018. PLoS One. 2018. PMID: 29995887 Free PMC article.
References
-
- Holliday R. The inheritance of epigenetic defects. Science. 1987; 238: 163–170 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
