

Horizontal Transmission of Cytosolic Sup35 Prions by Extracellular Vesicles
- PMID: 27406566
- PMCID: PMC4958257
- DOI: 10.1128/mBio.00915-16
Horizontal Transmission of Cytosolic Sup35 Prions by Extracellular Vesicles
Abstract
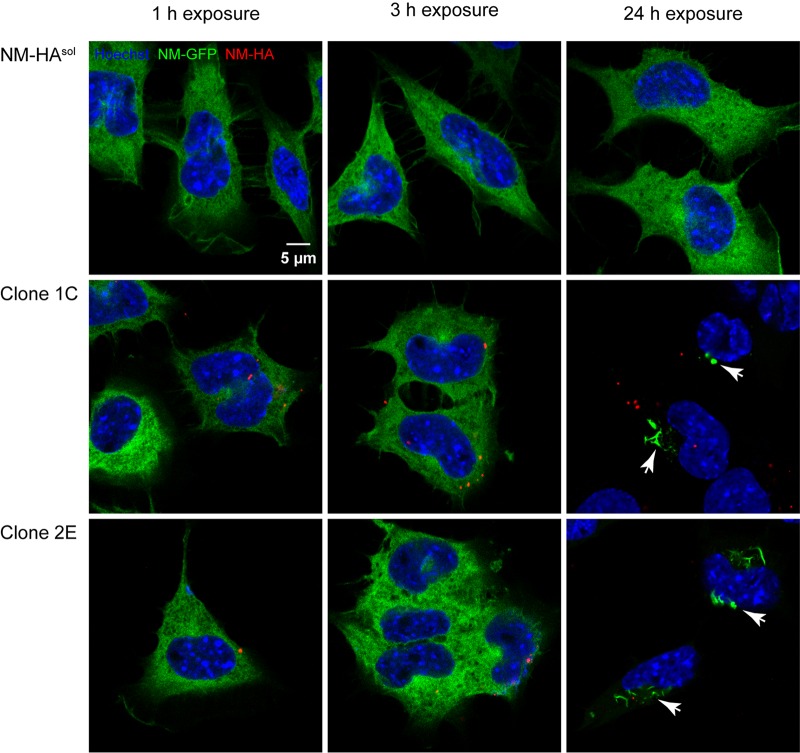
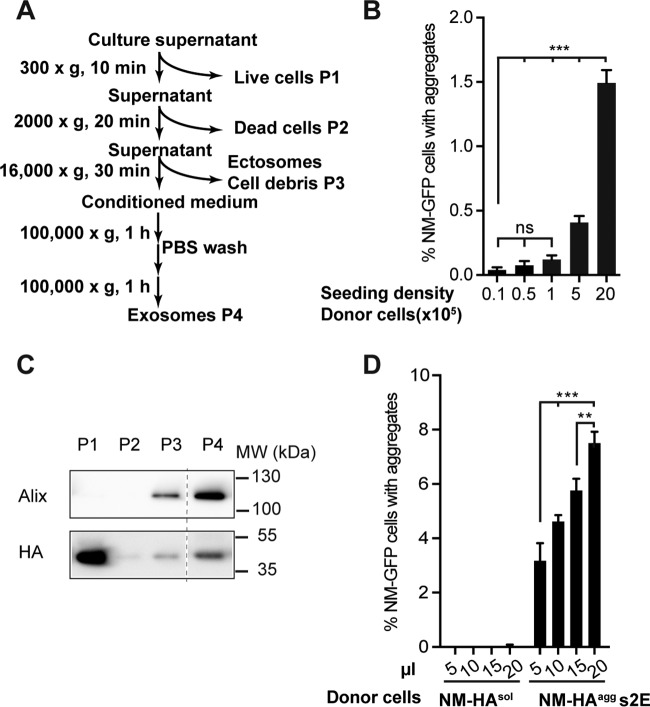
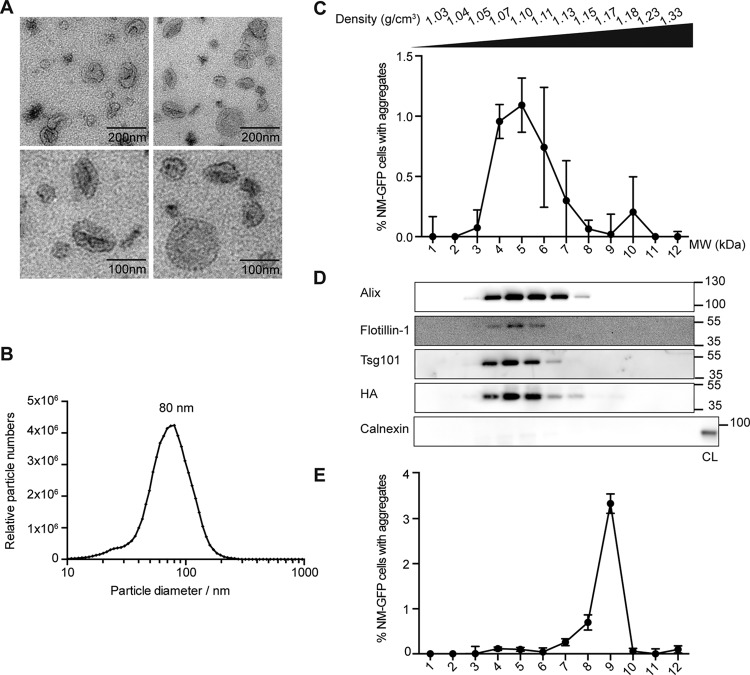
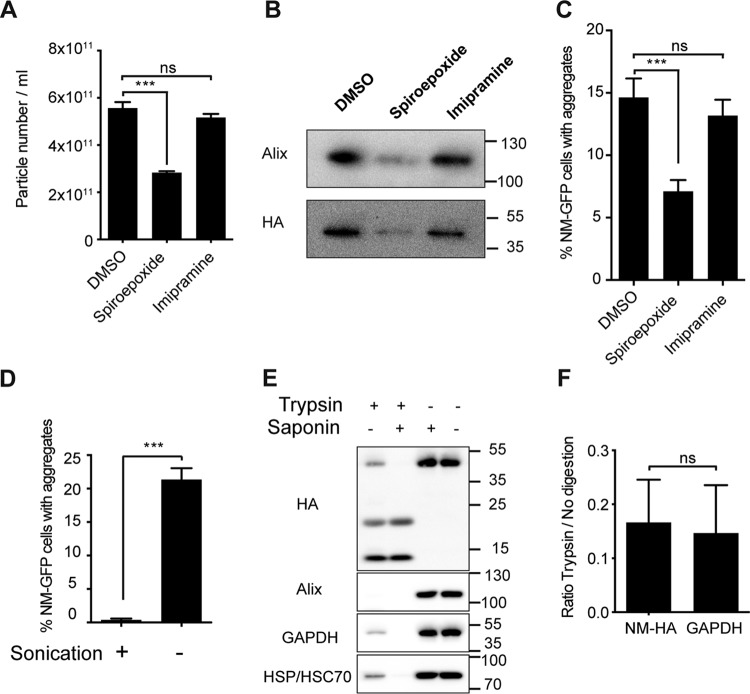
Prions are infectious protein particles that replicate by templating their aggregated state onto soluble protein of the same type. Originally identified as the causative agent of transmissible spongiform encephalopathies, prions in yeast (Saccharomyces cerevisiae) are epigenetic elements of inheritance that induce phenotypic changes of their host cells. The prototype yeast prion is the translation termination factor Sup35. Prions composed of Sup35 or its modular prion domain NM are heritable and are transmitted vertically to progeny or horizontally during mating. Interestingly, in mammalian cells, protein aggregates derived from yeast Sup35 NM behave as true infectious entities that employ dissemination strategies similar to those of mammalian prions. While transmission is most efficient when cells are in direct contact, we demonstrate here that cytosolic Sup35 NM prions are also released into the extracellular space in association with nanometer-sized membrane vesicles. Importantly, extracellular vesicles are biologically active and are taken up by recipient cells, where they induce self-sustained Sup35 NM protein aggregation. Thus, in mammalian cells, extracellular vesicles can serve as dissemination vehicles for protein-based epigenetic information transfer.
Importance: Prions are proteinaceous infectious particles that propagate by templating their quaternary structure onto nascent proteins of the same kind. Prions in yeast act as heritable epigenetic elements that can alter the phenotype when transmitted to daughter cells or during mating. Prion activity is conferred by so-called prion domains often enriched in glutamine and asparagine residues. Interestingly, many mammalian proteins also contain domains with compositional similarity to yeast prion domains. We have recently provided a proof-of-principle demonstration that a yeast prion domain also retains its prion activity in mammalian cells. We demonstrate here that cytosolic prions composed of a yeast prion domain are also packaged into extracellular vesicles that transmit the prion phenotype to bystander cells. Thus, proteins with prion-like domains can behave as proteinaceous information molecules that exploit the cellular vesicle trafficking machinery for intercellular long-distance dissemination.
Copyright © 2016 Liu et al.
Figures
References
-
- Moore RA, Vorberg I, Priola SA. 2005. Species barriers in prion diseases—brief review. Arch Virol Suppl 2005:187–202. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
