

Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons
- PMID: 27407143
- PMCID: PMC4968767
- DOI: 10.1073/pnas.1520387113
Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons
Abstract
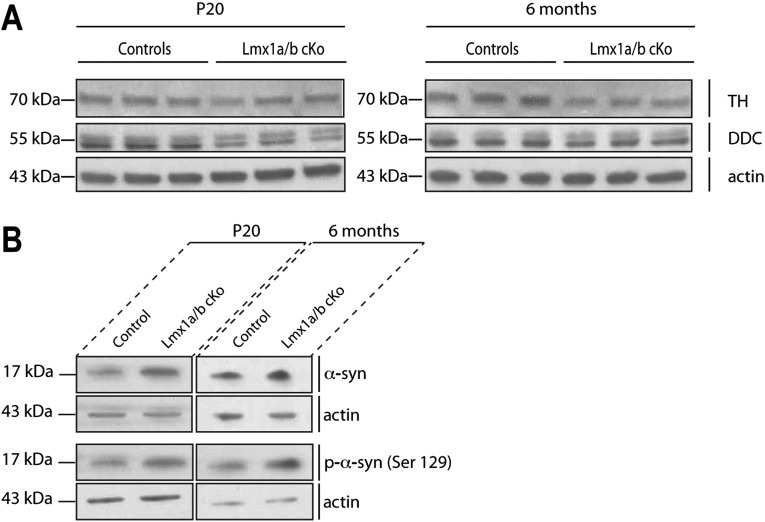
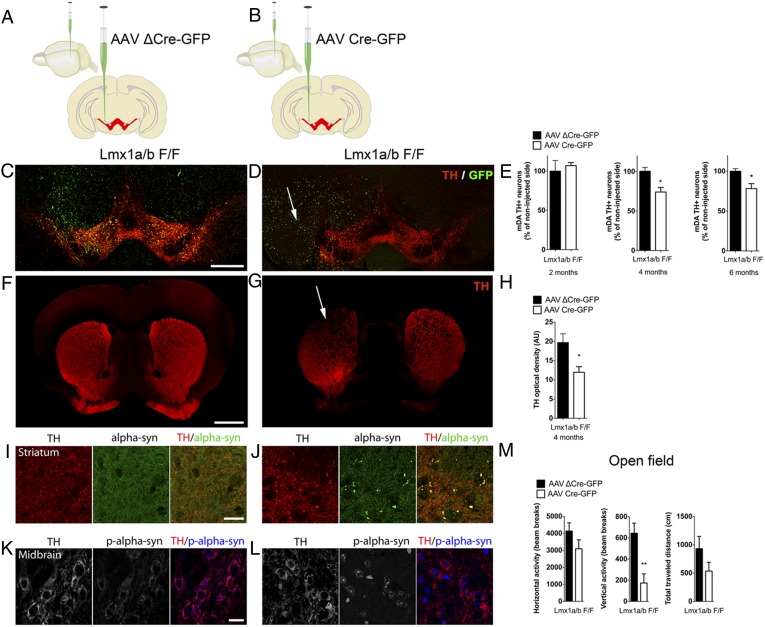
The LIM-homeodomain transcription factors Lmx1a and Lmx1b play critical roles during the development of midbrain dopaminergic progenitors, but their functions in the adult brain remain poorly understood. We show here that sustained expression of Lmx1a and Lmx1b is required for the survival of adult midbrain dopaminergic neurons. Strikingly, inactivation of Lmx1a and Lmx1b recreates cellular features observed in Parkinson's disease. We found that Lmx1a/b control the expression of key genes involved in mitochondrial functions, and their ablation results in impaired respiratory chain activity, increased oxidative stress, and mitochondrial DNA damage. Lmx1a/b deficiency caused axonal pathology characterized by α-synuclein(+) inclusions, followed by a progressive loss of dopaminergic neurons. These results reveal the key role of these transcription factors beyond the early developmental stages and provide mechanistic links between mitochondrial dysfunctions, α-synuclein aggregation, and the survival of dopaminergic neurons.
Keywords: Parkinson's disease; dopamine neurons; mitochondrial dysfunctions; protein aggregates; transcription factors.
Conflict of interest statement
The authors declare no conflict of interest.
Figures















References
-
- Shulman JM, De Jager PL, Feany MB. Parkinson’s disease: Genetics and pathogenesis. Annu Rev Pathol. 2011;6(1):193–222. - PubMed
-
- Abou-Sleiman PM, Muqit MM, Wood NW. Expanding insights of mitochondrial dysfunction in Parkinson’s disease. Nat Rev Neurosci. 2006;7(3):207–219. - PubMed
-
- Blaess S, Ang SL. Genetic control of midbrain dopaminergic neuron development. Wiley Interdiscip Rev Dev Biol. 2015;4(2):113–134. - PubMed
-
- Hegarty SV, Sullivan AM, O’Keeffe GW. Midbrain dopaminergic neurons: A review of the molecular circuitry that regulates their development. Dev Biol. 2013;379(2):123–138. - PubMed
-
- Doucet-Beaupré H, Lévesque M. 2013. The role of developmental transcription factors in adult midbrain dopaminergic neurons. OA Neurosciences 1(1):3.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
