

Branched-chain amino acid restriction in Zucker-fatty rats improves muscle insulin sensitivity by enhancing efficiency of fatty acid oxidation and acyl-glycine export
- PMID: 27408778
- PMCID: PMC4921791
- DOI: 10.1016/j.molmet.2016.04.006
Branched-chain amino acid restriction in Zucker-fatty rats improves muscle insulin sensitivity by enhancing efficiency of fatty acid oxidation and acyl-glycine export
Abstract
Objective: A branched-chain amino acid (BCAA)-related metabolic signature is strongly associated with insulin resistance and predictive of incident diabetes and intervention outcomes. To better understand the role that this metabolite cluster plays in obesity-related metabolic dysfunction, we studied the impact of BCAA restriction in a rodent model of obesity in which BCAA metabolism is perturbed in ways that mirror the human condition.
Methods: Zucker-lean rats (ZLR) and Zucker-fatty rats (ZFR) were fed either a custom control, low fat (LF) diet, or an isonitrogenous, isocaloric LF diet in which all three BCAA (Leu, Ile, Val) were reduced by 45% (LF-RES). We performed comprehensive metabolic and physiologic profiling to characterize the effects of BCAA restriction on energy balance, insulin sensitivity, and glucose, lipid and amino acid metabolism.
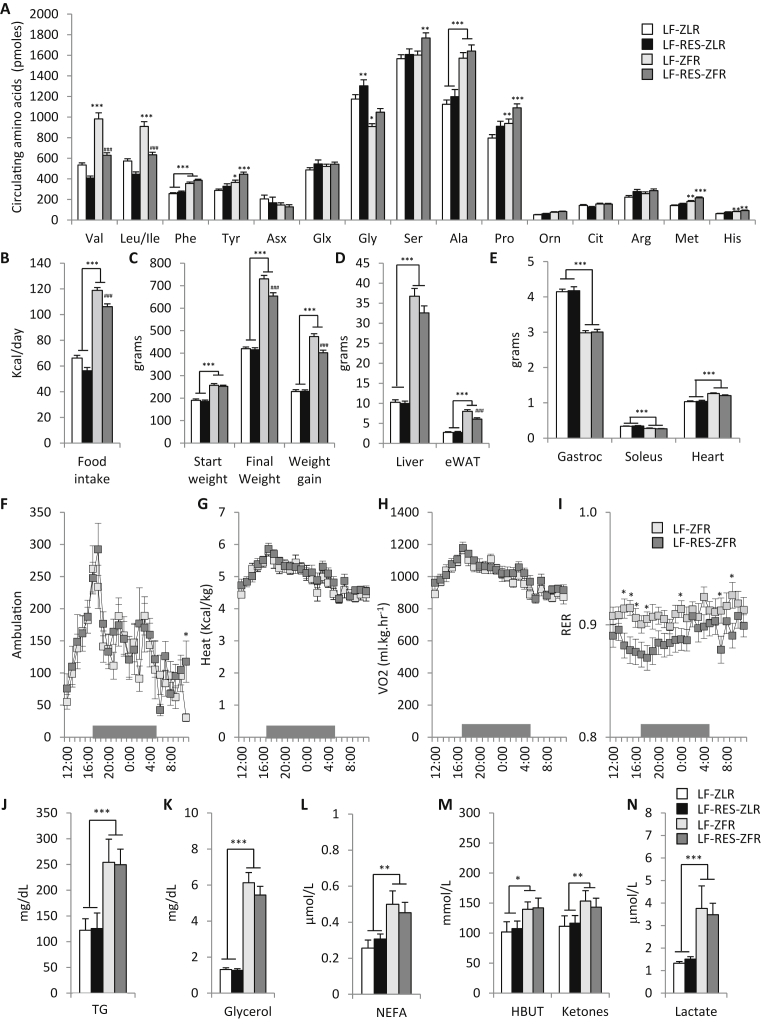
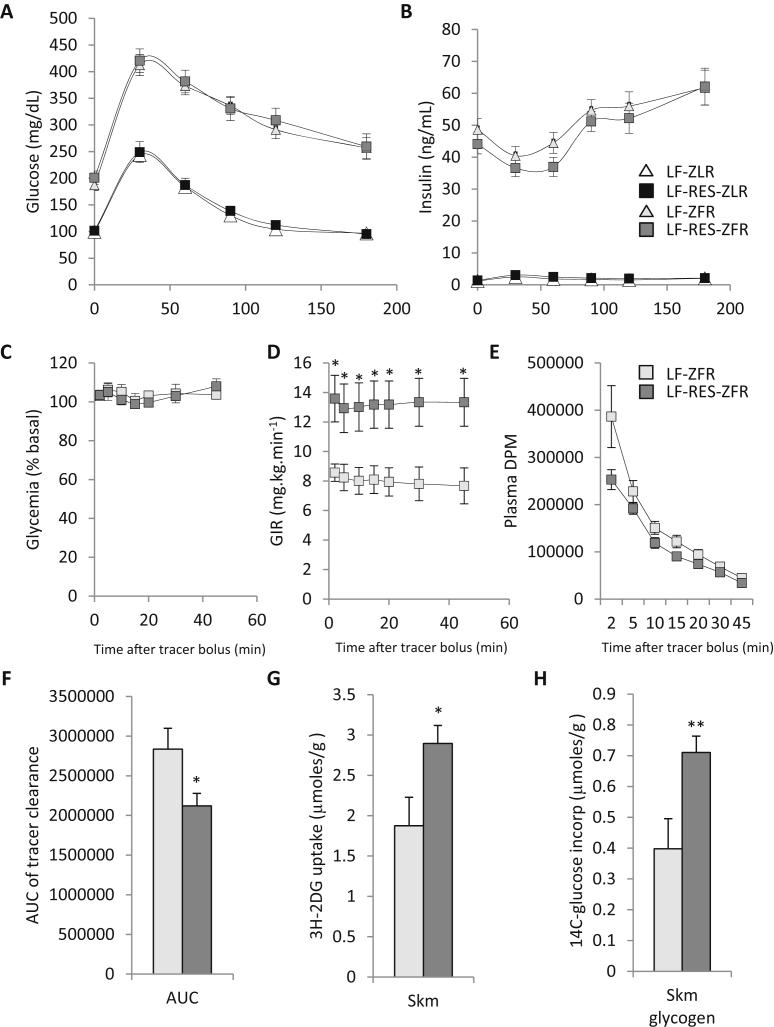
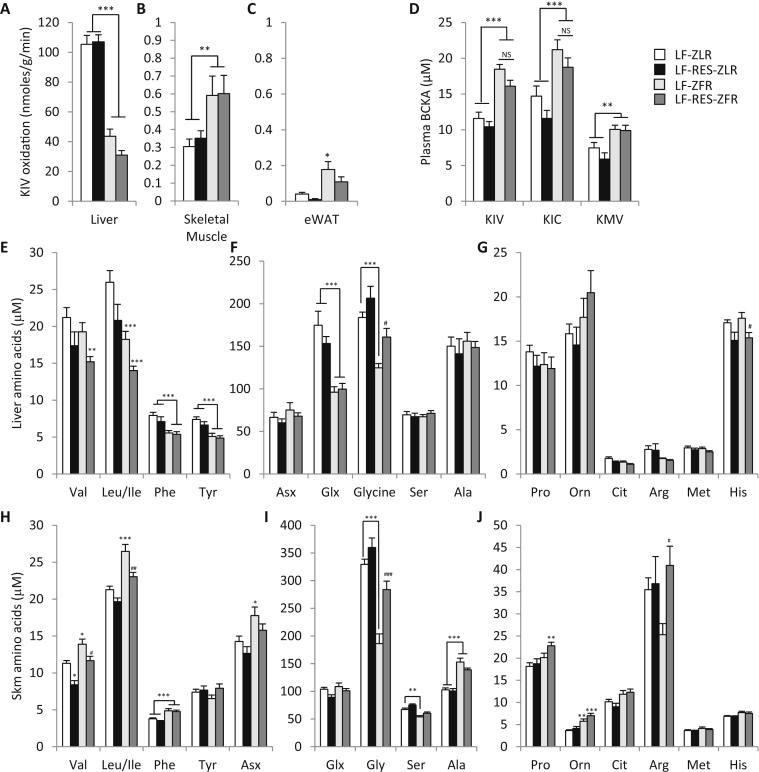
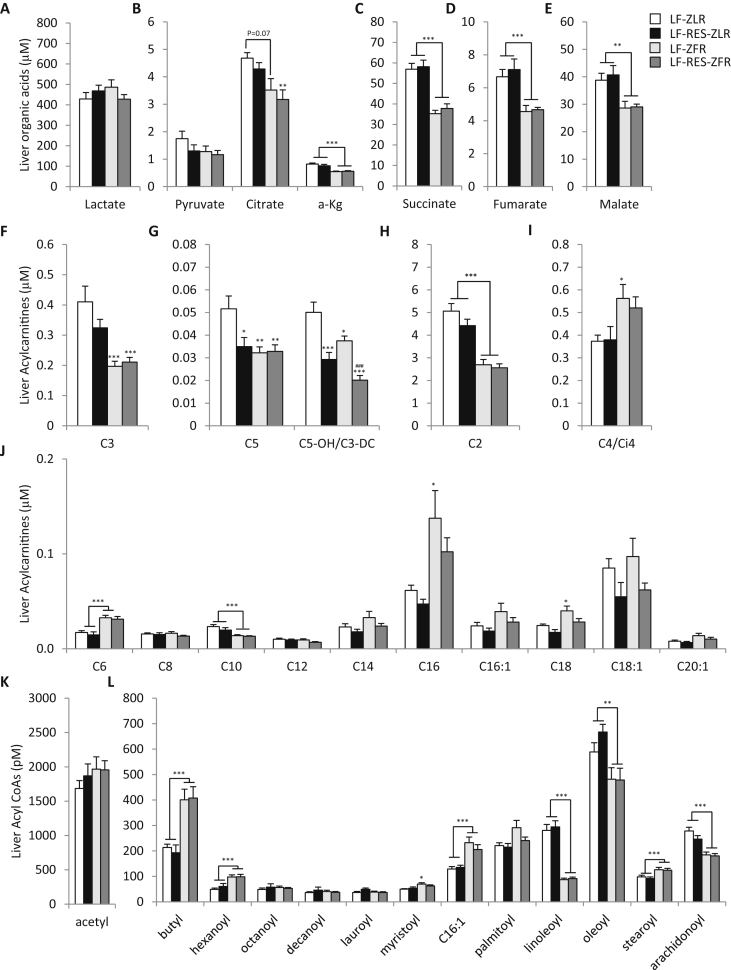
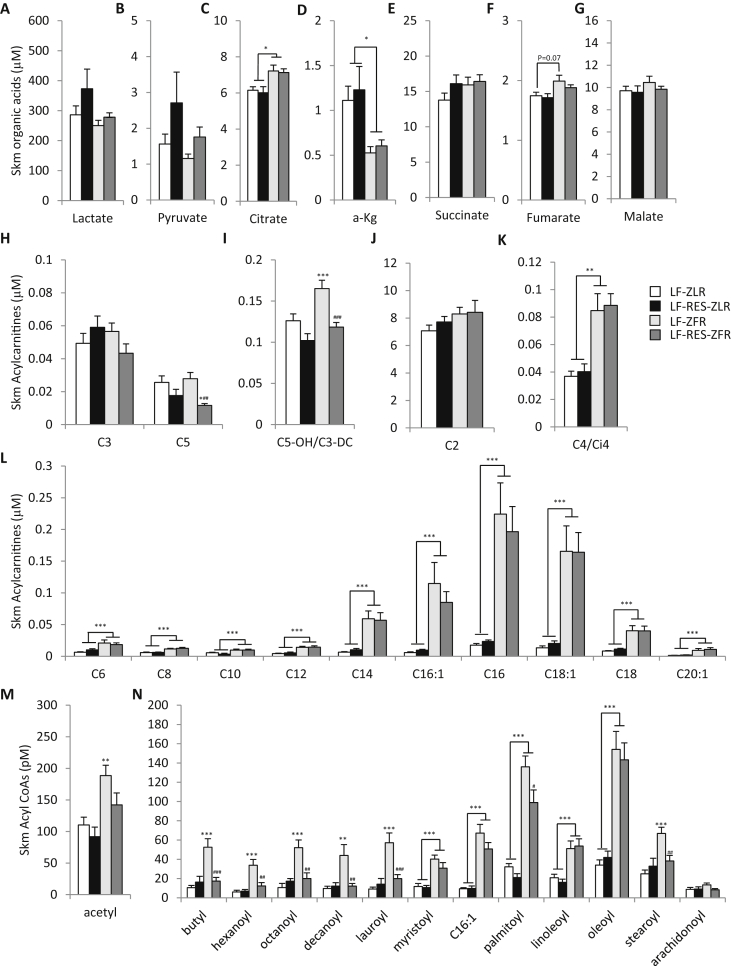
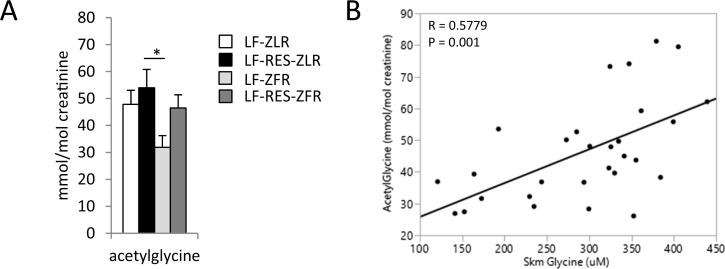
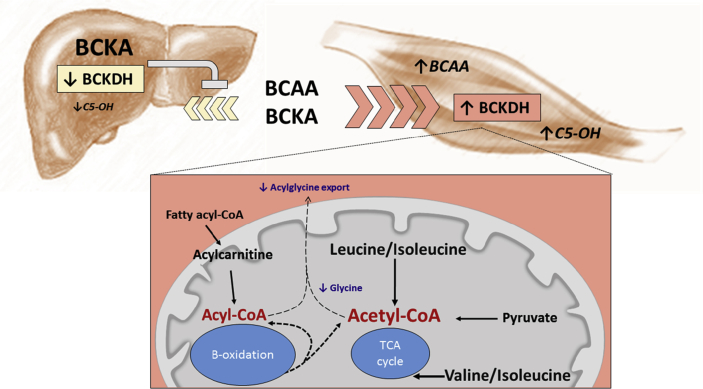
Results: LF-fed ZFR had higher levels of circulating BCAA and lower levels of glycine compared to LF-fed ZLR. Feeding ZFR with the LF-RES diet lowered circulating BCAA to levels found in LF-fed ZLR. Activity of the rate limiting enzyme in the BCAA catabolic pathway, branched chain keto acid dehydrogenase (BCKDH), was lower in liver but higher in skeletal muscle of ZFR compared to ZLR and was not responsive to diet in either tissue. BCAA restriction had very little impact on metabolites studied in liver of ZFR where BCAA content was low, and BCKDH activity was suppressed. However, in skeletal muscle of LF-fed ZFR compared to LF-fed ZLR, where BCAA content and BCKDH activity were increased, accumulation of fatty acyl CoAs was completely normalized by dietary BCAA restriction. BCAA restriction also normalized skeletal muscle glycine content and increased urinary acetyl glycine excretion in ZFR. These effects were accompanied by lower RER and improved skeletal muscle insulin sensitivity in LF-RES fed ZFR as measured by hyperinsulinemic-isoglycemic clamp.
Conclusions: Our data are consistent with a model wherein elevated circulating BCAA contribute to development of obesity-related insulin resistance by interfering with lipid oxidation in skeletal muscle. BCAA-dependent lowering of the skeletal muscle glycine pool appears to contribute to this effect by slowing acyl-glycine export to the urine.
Keywords: BCAA; Insulin sensitivity; Metabolism; Obesity.
Figures
References
-
- Felig P., Marliss E., Cahill G.F. Plasma amino acid levels and insulin secretion in obesity. The New England Journal of Medicine. 1969;281(15):811–816. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
