

The Actin Filament-Binding Protein Coronin Regulates Motility in Plasmodium Sporozoites
- PMID: 27409081
- PMCID: PMC4943629
- DOI: 10.1371/journal.ppat.1005710
The Actin Filament-Binding Protein Coronin Regulates Motility in Plasmodium Sporozoites
Erratum in
-
Correction: The Actin Filament-Binding Protein Coronin Regulates Motility in Plasmodium Sporozoites.PLoS Pathog. 2016 Aug 2;12(8):e1005818. doi: 10.1371/journal.ppat.1005818. eCollection 2016 Aug. PLoS Pathog. 2016. PMID: 27482908 Free PMC article.
Abstract
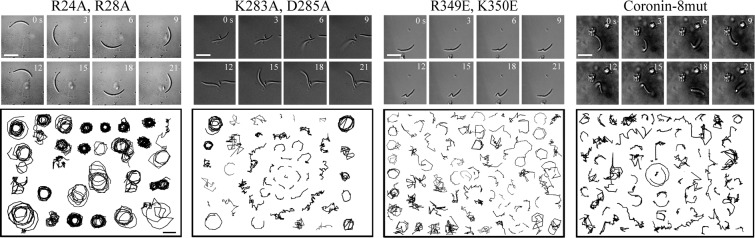
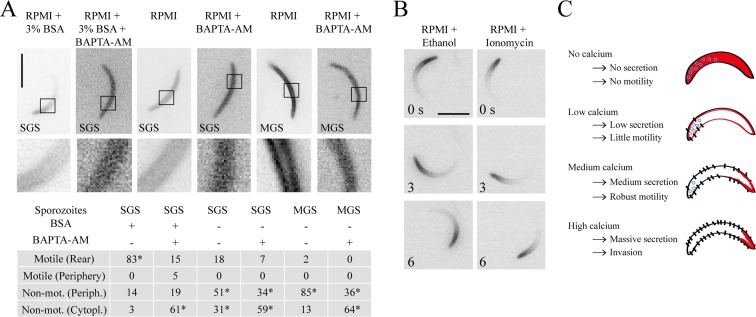
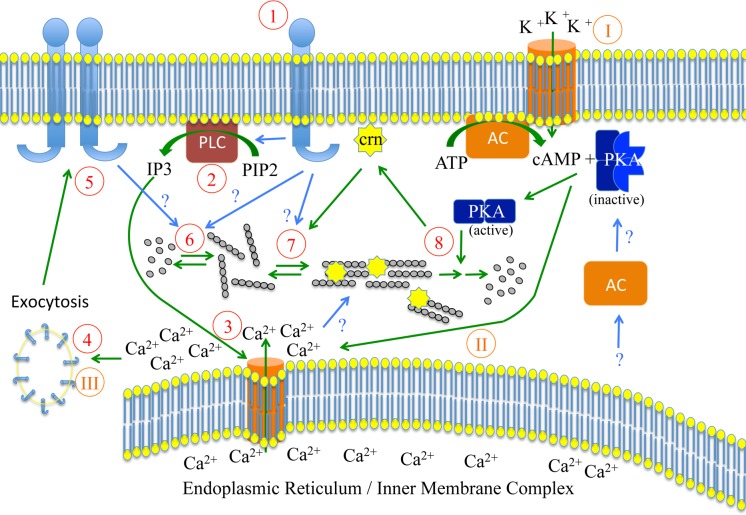
Parasites causing malaria need to migrate in order to penetrate tissue barriers and enter host cells. Here we show that the actin filament-binding protein coronin regulates gliding motility in Plasmodium berghei sporozoites, the highly motile forms of a rodent malaria-causing parasite transmitted by mosquitoes. Parasites lacking coronin show motility defects that impair colonization of the mosquito salivary glands but not migration in the skin, yet result in decreased transmission efficiency. In non-motile sporozoites low calcium concentrations mediate actin-independent coronin localization to the periphery. Engagement of extracellular ligands triggers an intracellular calcium release followed by the actin-dependent relocalization of coronin to the rear and initiation of motility. Mutational analysis and imaging suggest that coronin organizes actin filaments for productive motility. Using coronin-mCherry as a marker for the presence of actin filaments we found that protein kinase A contributes to actin filament disassembly. We finally speculate that calcium and cAMP-mediated signaling regulate a switch from rapid parasite motility to host cell invasion by differentially influencing actin dynamics.
Conflict of interest statement
I have read the journal's policy and the authors of this manuscript have the following competing interests: KH is shareholder of MalVa GmbH, a spin-off company of the Heidelberg University Hospital focussing on malaria vaccine development. There are no patents, products in development, marketed products or financial interests to declare. This does not alter our adherence to all PLOS Pathogens policies on sharing data and materials.
Figures



References
-
- Vanderberg JP, Frevert U (2004) Intravital microscopy demonstrating antibody-mediated immobilisation of Plasmodium berghei sporozoites injected into skin by mosquitoes. Int J Parasitol 34: 991–996. - PubMed
-
- Amino R, Thiberge S, Martin B, Celli S, Shorte S, et al. (2006) Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med 12: 220–224. - PubMed
-
- Prudencio M, Rodriguez A, Mota MM (2006) The silent path to thousands of merozoites: the Plasmodium liver stage. Nat Rev Microbiol 4: 849–856. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
