

Hypoxia-mediated alterations and their role in the HER-2/neuregulated CREB status and localization
- PMID: 27409833
- PMCID: PMC5239535
- DOI: 10.18632/oncotarget.10474
Hypoxia-mediated alterations and their role in the HER-2/neuregulated CREB status and localization
Abstract
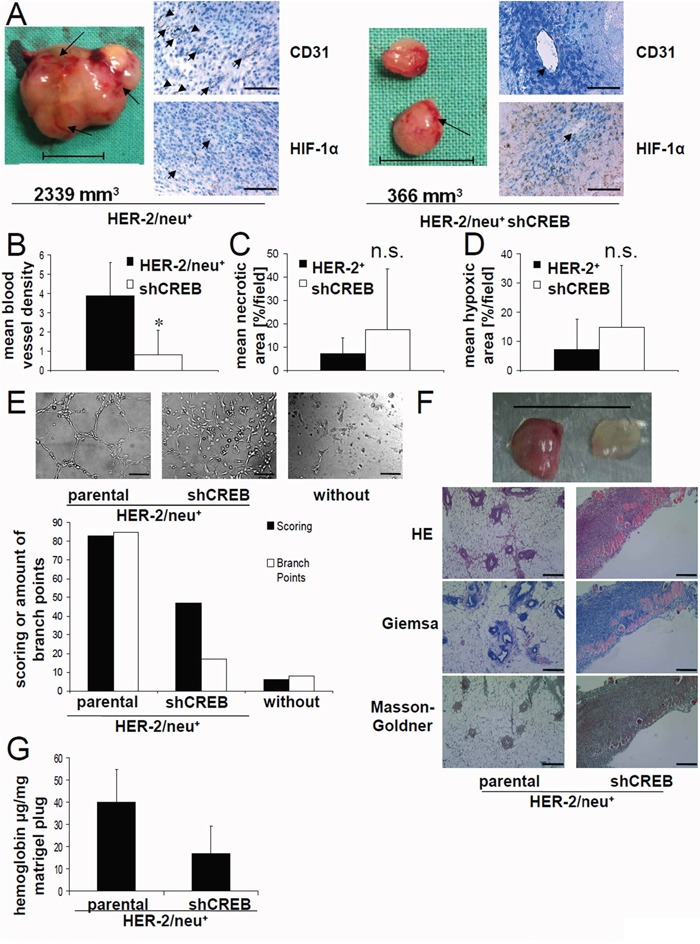
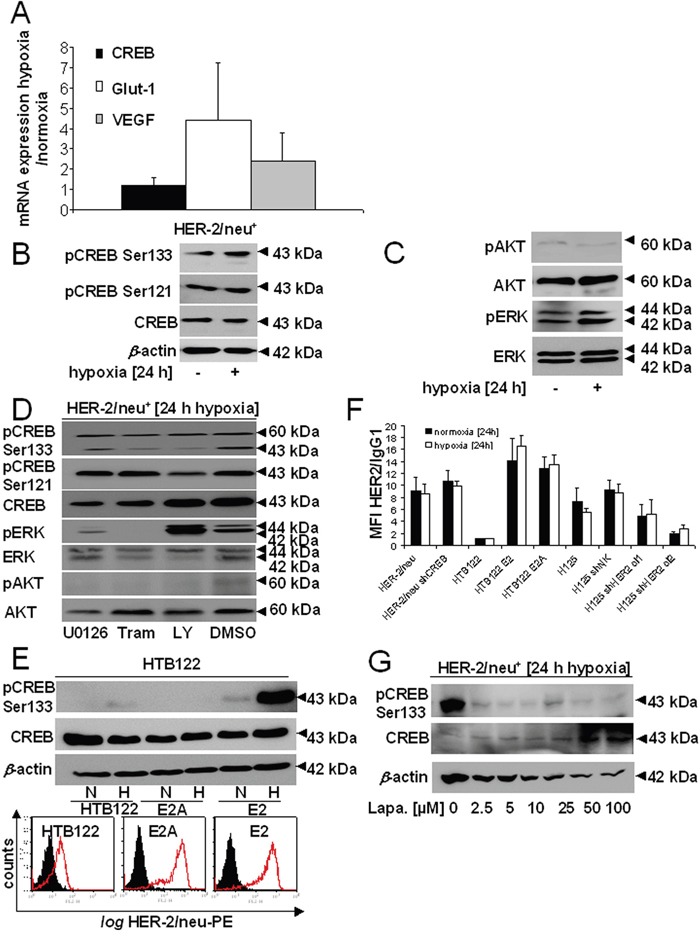
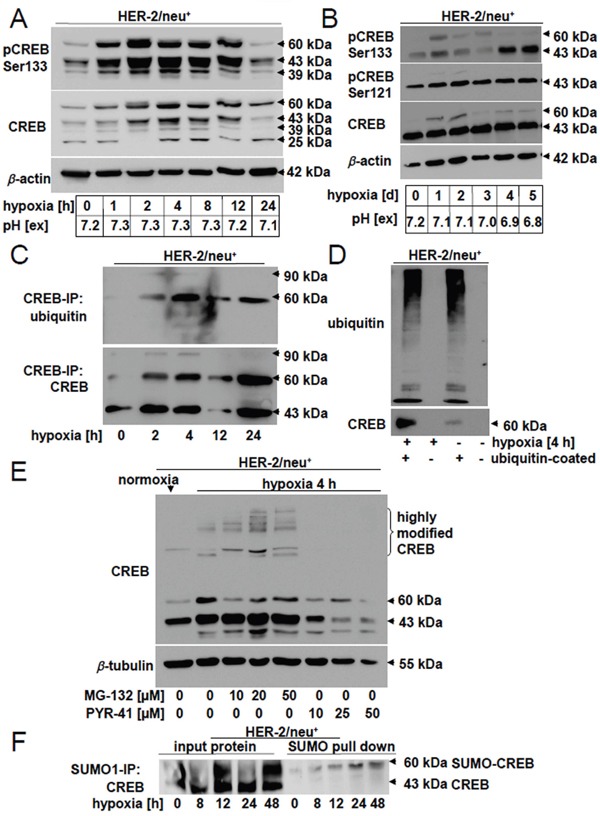
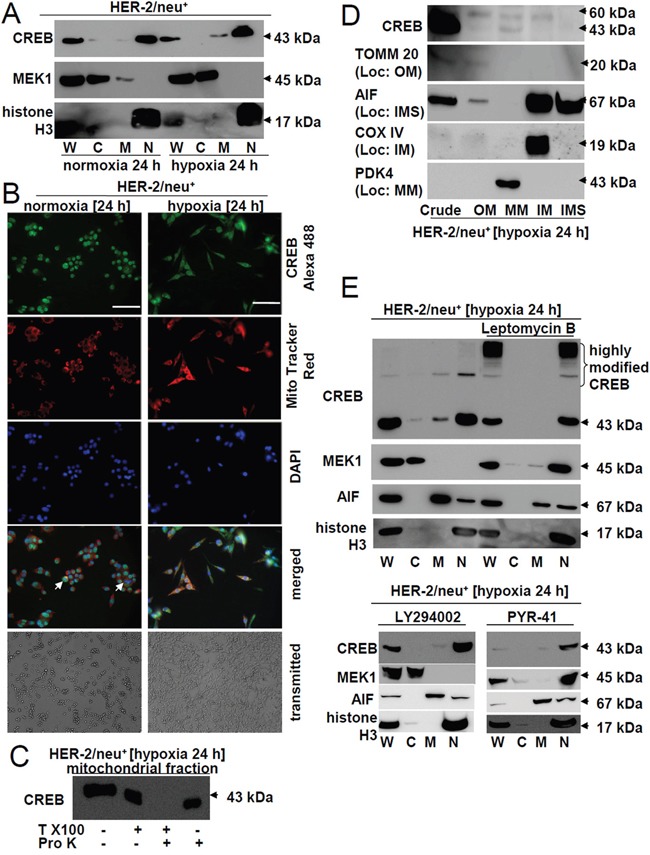
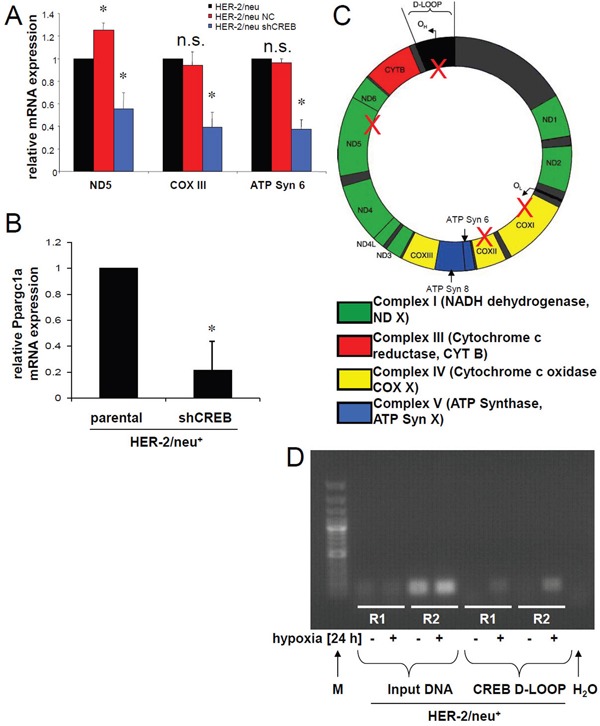
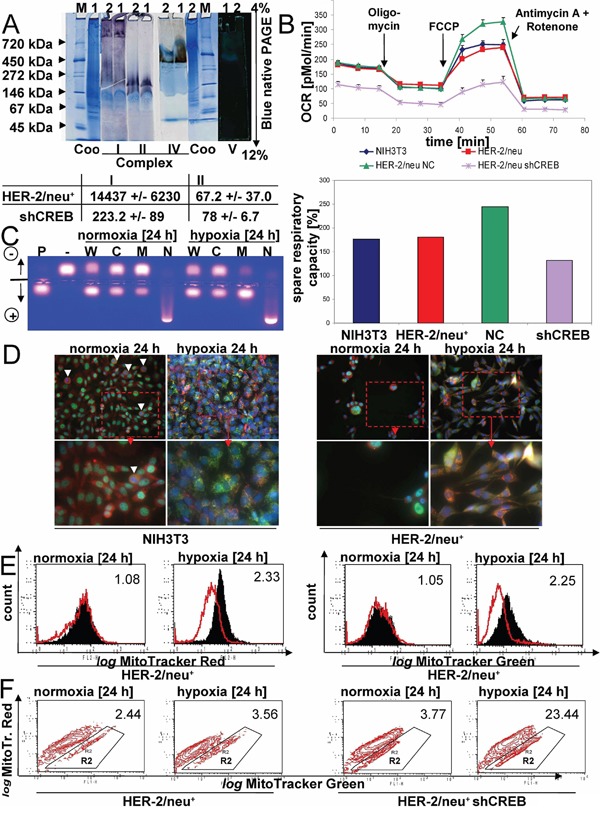
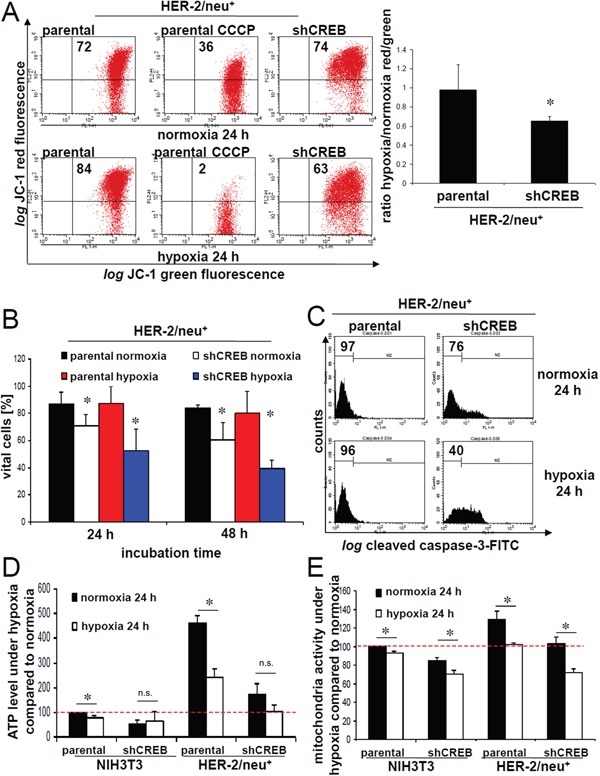
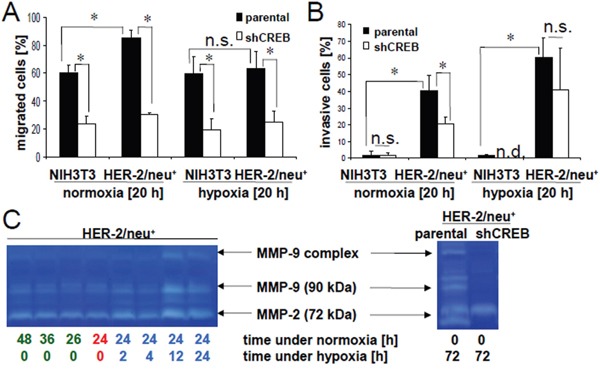
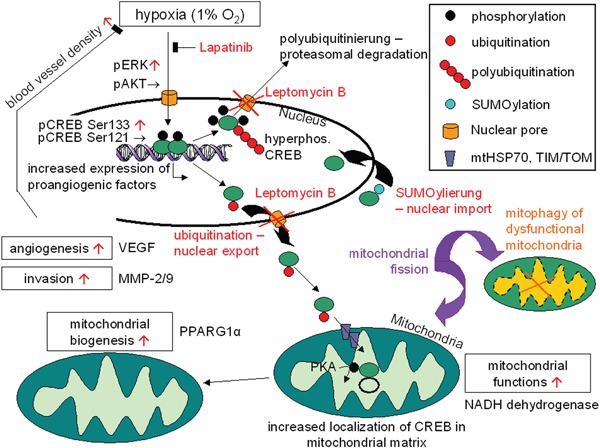
The cAMP-responsive element-binding protein (CREB) is involved in the tumorigenicity of HER-2/neu-overexpressing murine and human tumor cells, but a link between the HER-2/neu-mediated CREB activation, its posttranslational modification and localization and changes in the cellular metabolism, due to an altered (tumor) microenvironment remains to be established. The present study demonstrated that shRNA-mediated silencing of CREB in HER-2/neu-transformed cells resulted in decreased tumor formation, which was associated with reduced angiogenesis, but increased necrotic and hypoxic areas in the tumor. Hypoxia induced pCREBSer133, but not pCREBSer121 expression in HER-2/neu-transformed cells. This was accompanied by upregulation of the hypoxia-inducible genes GLUT1 and VEGF, increased cell migration and matrix metalloproteinase-mediated invasion. Treatment of HER-2/neu+ cells with signal transduction inhibitors targeting in particular HER-2/neu was able to revert hypoxia-controlled CREB activation. In addition to changes in the phosphorylation, hypoxic response of HER-2/neu+ cells caused a transient ubiquitination and SUMOylation as well as a co-localization of nuclear CREB to the mitochondrial matrix. A mitochondrial localization of CREB was also demonstrated in hypoxic areas of HER-2/neu+ mammary carcinoma lesions. This was accompanied by an altered gene expression pattern, activity and metabolism of mitochondria leading to an increased respiratory rate, oxidative phosphorylation and mitochondrial membrane potential and consequently to an enhanced apoptosis and reduced cell viability. These data suggest that the HER-2/neu-mediated CREB activation caused by a hypoxic tumor microenvironment contributes to the neoplastic phenotype of HER-2/neu+ cells at various levels.
Keywords: CREB; HER-2/neu; angiogenesis; hypoxia; mitochondria.
Conflict of interest statement
The authors declare to have no conflicts of interest.
Figures
Similar articles
-
Linking CREB function with altered metabolism in murine fibroblast-based model cell lines.Oncotarget. 2017 Oct 27;8(57):97439-97463. doi: 10.18632/oncotarget.22135. eCollection 2017 Nov 14. Oncotarget. 2017. PMID: 29228623 Free PMC article.
-
HER-2/neu mediates oncogenic transformation via altered CREB expression and function.Mol Cancer Res. 2013 Nov;11(11):1462-77. doi: 10.1158/1541-7786.MCR-13-0125. Epub 2013 Sep 11. Mol Cancer Res. 2013. PMID: 24025972
-
Colorectal Carcinogenesis: Connecting K-RAS-Induced Transformation and CREB Activity In Vitro and In Vivo.Mol Cancer Res. 2015 Aug;13(8):1248-62. doi: 10.1158/1541-7786.MCR-14-0590. Epub 2015 May 1. Mol Cancer Res. 2015. PMID: 25934695
-
HER-2/neu-mediated down-regulation of biglycan associated with altered growth properties.J Biol Chem. 2012 Jul 13;287(29):24320-9. doi: 10.1074/jbc.M111.334425. Epub 2012 May 11. J Biol Chem. 2012. PMID: 22582394 Free PMC article.
-
Protein tyrosine phosphatase epsilon and Neu-induced mammary tumorigenesis.Cancer Metastasis Rev. 2008 Jun;27(2):193-203. doi: 10.1007/s10555-008-9124-0. Cancer Metastasis Rev. 2008. PMID: 18231724 Review.
Cited by
-
Hypoxia Induced Sex-Difference in Zebrafish Brain Proteome Profile Reveals the Crucial Role of H3K9me3 in Recovery From Acute Hypoxia.Front Genet. 2022 Jan 31;12:635904. doi: 10.3389/fgene.2021.635904. eCollection 2021. Front Genet. 2022. PMID: 35173759 Free PMC article.
-
Biglycan-mediated upregulation of MHC class I expression in HER-2/neu-transformed cells.Oncoimmunology. 2018 Jan 16;7(4):e1373233. doi: 10.1080/2162402X.2017.1373233. eCollection 2018. Oncoimmunology. 2018. PMID: 29632715 Free PMC article.
-
IGFBP5 is an ROR1 ligand promoting glioblastoma invasion via ROR1/HER2-CREB signaling axis.Nat Commun. 2023 Mar 22;14(1):1578. doi: 10.1038/s41467-023-37306-1. Nat Commun. 2023. PMID: 36949068 Free PMC article.
-
What turns CREB on? And off? And why does it matter?Cell Mol Life Sci. 2020 Oct;77(20):4049-4067. doi: 10.1007/s00018-020-03525-8. Epub 2020 Apr 28. Cell Mol Life Sci. 2020. PMID: 32347317 Free PMC article. Review.
-
Linking CREB function with altered metabolism in murine fibroblast-based model cell lines.Oncotarget. 2017 Oct 27;8(57):97439-97463. doi: 10.18632/oncotarget.22135. eCollection 2017 Nov 14. Oncotarget. 2017. PMID: 29228623 Free PMC article.
References
-
- Dodson GE, Tibbetts RS. DNA replication stress-induced phosphorylation of cyclic AMP response element-binding protein mediated by ATM. J Biol Chem. 2006;281:1692–1697. - PubMed
-
- Johannessen M, Delghandi MP, Moens U. What turns CREB on? Cell Signal. 2004;16:1211–1227. - PubMed
-
- Ishimoto T, Mano H, Ozawa T, Mori H. Measuring CREB activation using bioluminescent probes that detect KID-KIX interaction in living cells. Bioconjug Chem. 2012;23:923–932. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
