

LHP1 Regulates H3K27me3 Spreading and Shapes the Three-Dimensional Conformation of the Arabidopsis Genome
- PMID: 27410265
- PMCID: PMC4943711
- DOI: 10.1371/journal.pone.0158936
LHP1 Regulates H3K27me3 Spreading and Shapes the Three-Dimensional Conformation of the Arabidopsis Genome
Abstract
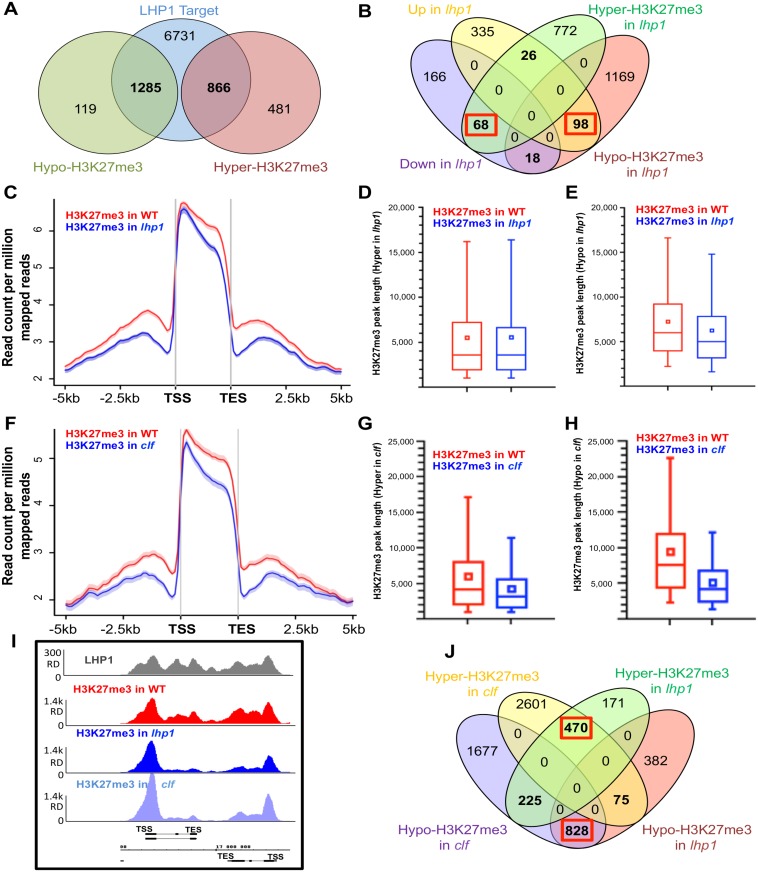
Precise expression patterns of genes in time and space are essential for proper development of multicellular organisms. Dynamic chromatin conformation and spatial organization of the genome constitute a major step in this regulation to modulate developmental outputs. Polycomb repressive complexes (PRCs) mediate stable or flexible gene repression in response to internal and environmental cues. In Arabidopsis thaliana, LHP1 co-localizes with H3K27me3 epigenetic marks throughout the genome and interacts with PRC1 and PRC2 members as well as with a long noncoding RNA. Here, we show that LHP1 is responsible for the spreading of H3K27me3 towards the 3' end of the gene body. We also identified a subset of LHP1-activated genes and demonstrated that LHP1 shapes local chromatin topology in order to control transcriptional co-regulation. Our work reveals a general role of LHP1 from local to higher conformation levels of chromatin configuration to determine its accessibility to define gene expression patterns.
Conflict of interest statement
Figures




Similar articles
-
H2A monoubiquitination in Arabidopsis thaliana is generally independent of LHP1 and PRC2 activity.Genome Biol. 2017 Apr 12;18(1):69. doi: 10.1186/s13059-017-1197-z. Genome Biol. 2017. PMID: 28403905 Free PMC article.
-
Transcription factors AS1 and AS2 interact with LHP1 to repress KNOX genes in Arabidopsis.J Integr Plant Biol. 2016 Dec;58(12):959-970. doi: 10.1111/jipb.12485. Epub 2016 Jul 20. J Integr Plant Biol. 2016. PMID: 27273574
-
Ctf4-related protein recruits LHP1-PRC2 to maintain H3K27me3 levels in dividing cells in Arabidopsis thaliana.Proc Natl Acad Sci U S A. 2017 May 2;114(18):4833-4838. doi: 10.1073/pnas.1620955114. Epub 2017 Apr 20. Proc Natl Acad Sci U S A. 2017. PMID: 28428341 Free PMC article.
-
Current understanding of plant Polycomb group proteins and the repressive histone H3 Lysine 27 trimethylation.Biochem Soc Trans. 2020 Aug 28;48(4):1697-1706. doi: 10.1042/BST20200192. Biochem Soc Trans. 2020. PMID: 32725200 Review.
-
Chromatin modulation and gene regulation in plants: insight about PRC1 function.Biochem Soc Trans. 2018 Aug 20;46(4):957-966. doi: 10.1042/BST20170576. Epub 2018 Jul 31. Biochem Soc Trans. 2018. PMID: 30065110 Review.
Cited by
-
HSI2/VAL1 and HSL1/VAL2 function redundantly to repress DOG1 expression in Arabidopsis seeds and seedlings.New Phytol. 2020 Aug;227(3):840-856. doi: 10.1111/nph.16559. Epub 2020 Apr 25. New Phytol. 2020. PMID: 32201955 Free PMC article.
-
Emerging roles of chromatin in the maintenance of genome organization and function in plants.Genome Biol. 2017 May 23;18(1):96. doi: 10.1186/s13059-017-1236-9. Genome Biol. 2017. PMID: 28535770 Free PMC article. Review.
-
The Impact of Polycomb Group Proteins on 3D Chromatin Structure and Environmental Stresses in Plants.Plants (Basel). 2025 Mar 27;14(7):1038. doi: 10.3390/plants14071038. Plants (Basel). 2025. PMID: 40219106 Free PMC article. Review.
-
PCSD: a plant chromatin state database.Nucleic Acids Res. 2018 Jan 4;46(D1):D1157-D1167. doi: 10.1093/nar/gkx919. Nucleic Acids Res. 2018. PMID: 29040761 Free PMC article.
-
Get closer and make hotspots: liquid-liquid phase separation in plants.EMBO Rep. 2021 May 5;22(5):e51656. doi: 10.15252/embr.202051656. Epub 2021 Apr 28. EMBO Rep. 2021. PMID: 33913240 Free PMC article. Review.
References
-
- Lewis EB. A gene complex controlling segmentation in Drosophila. Nature. 1978;276(5688):565–70. . - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
