

Microbiota Dysbiosis Controls the Neuroinflammatory Response after Stroke
- PMID: 27413153
- PMCID: PMC6705544
- DOI: 10.1523/JNEUROSCI.1114-16.2016
Microbiota Dysbiosis Controls the Neuroinflammatory Response after Stroke
Abstract
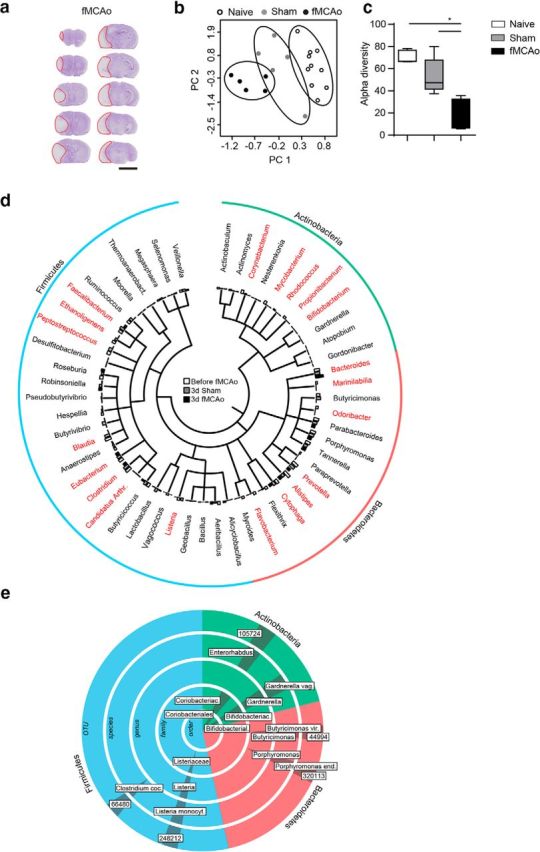
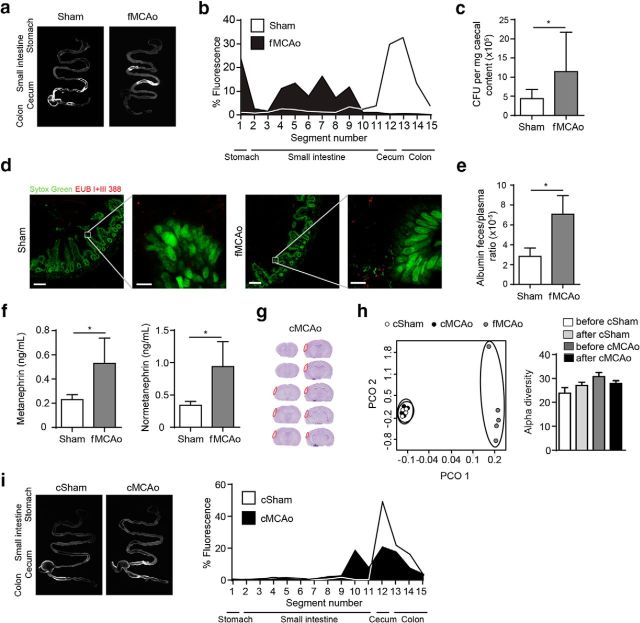
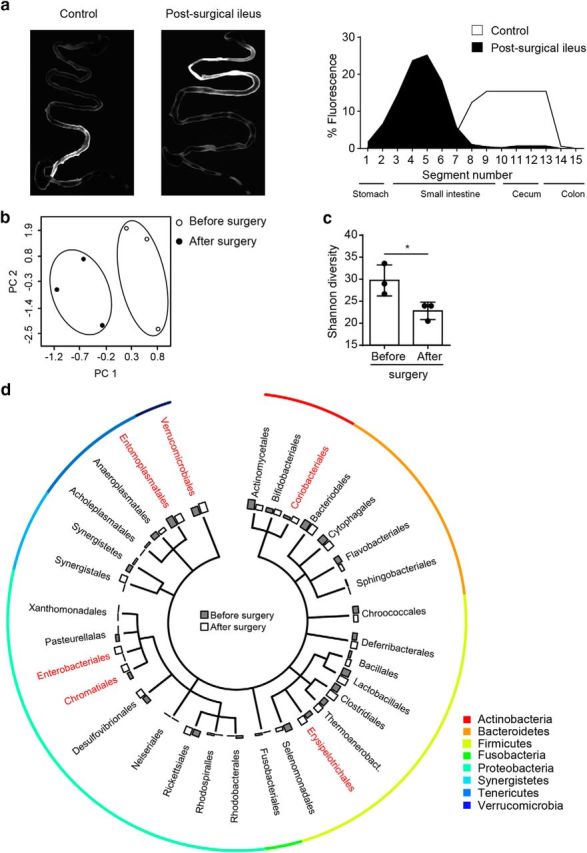
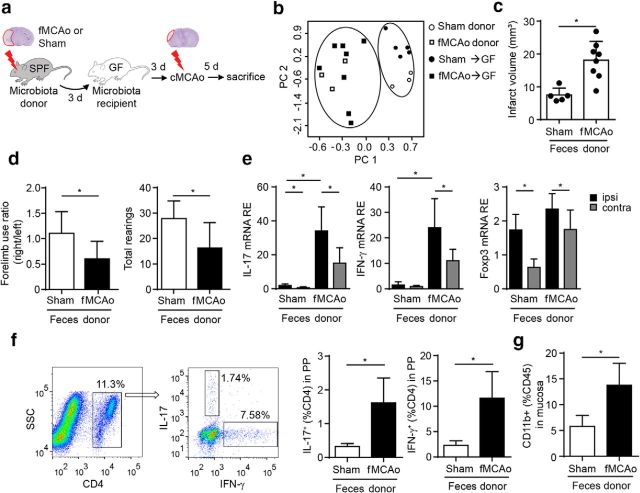
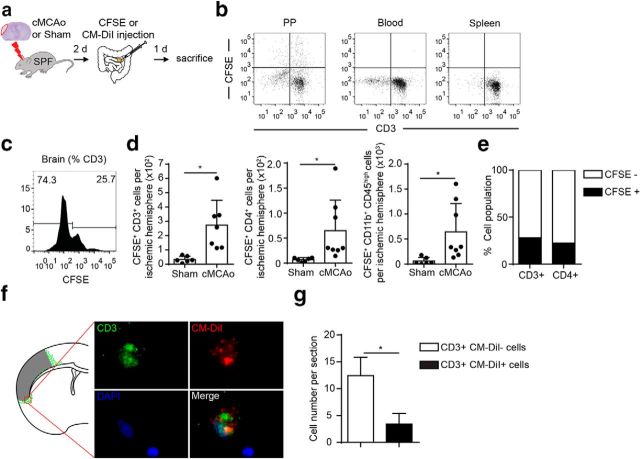
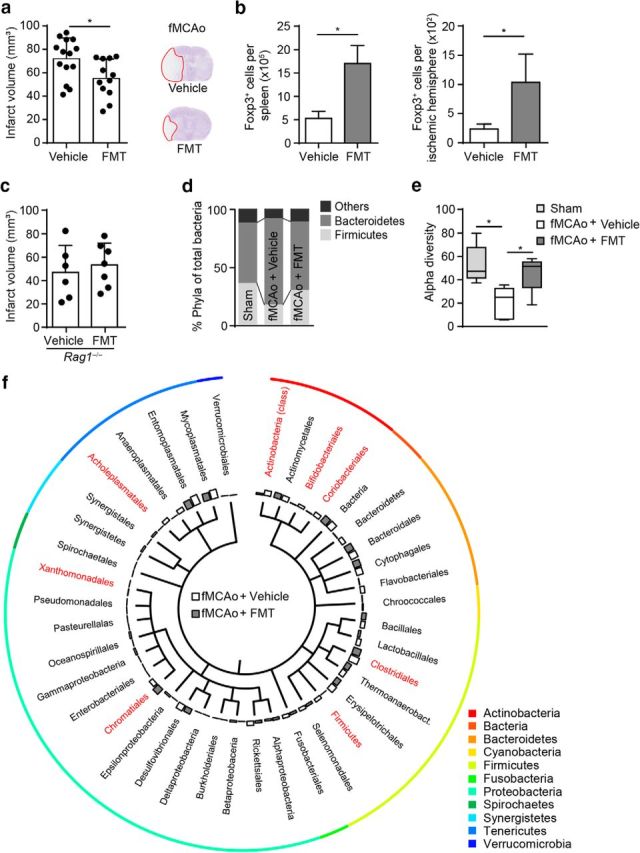
Acute brain ischemia induces a local neuroinflammatory reaction and alters peripheral immune homeostasis at the same time. Recent evidence has suggested a key role of the gut microbiota in autoimmune diseases by modulating immune homeostasis. Therefore, we investigated the mechanistic link among acute brain ischemia, microbiota alterations, and the immune response after brain injury. Using two distinct models of acute middle cerebral artery occlusion, we show by next-generation sequencing that large stroke lesions cause gut microbiota dysbiosis, which in turn affects stroke outcome via immune-mediated mechanisms. Reduced species diversity and bacterial overgrowth of bacteroidetes were identified as hallmarks of poststroke dysbiosis, which was associated with intestinal barrier dysfunction and reduced intestinal motility as determined by in vivo intestinal bolus tracking. Recolonizing germ-free mice with dysbiotic poststroke microbiota exacerbates lesion volume and functional deficits after experimental stroke compared with the recolonization with a normal control microbiota. In addition, recolonization of mice with a dysbiotic microbiome induces a proinflammatory T-cell polarization in the intestinal immune compartment and in the ischemic brain. Using in vivo cell-tracking studies, we demonstrate the migration of intestinal lymphocytes to the ischemic brain. Therapeutic transplantation of fecal microbiota normalizes brain lesion-induced dysbiosis and improves stroke outcome. These results support a novel mechanism in which the gut microbiome is a target of stroke-induced systemic alterations and an effector with substantial impact on stroke outcome.
Significance statement: We have identified a bidirectional communication along the brain-gut microbiota-immune axis and show that the gut microbiota is a central regulator of immune homeostasis. Acute brain lesions induced dysbiosis of the microbiome and, in turn, changes in the gut microbiota affected neuroinflammatory and functional outcome after brain injury. The microbiota impact on immunity and stroke outcome was transmissible by microbiota transplantation. Our findings support an emerging concept in which the gut microbiota is a key regulator in priming the neuroinflammatory response to brain injury. These findings highlight the key role of microbiota as a potential therapeutic target to protect brain function after injury.
Keywords: T cells; inflammation; microbiota; stroke.
Copyright © 2016 the authors 0270-6474/16/367428-13$15.00/0.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical