

Genome-Wide Analysis of the bZIP Gene Family Identifies Two ABI5-Like bZIP Transcription Factors, BrABI5a and BrABI5b, as Positive Modulators of ABA Signalling in Chinese Cabbage
- PMID: 27414644
- PMCID: PMC4944949
- DOI: 10.1371/journal.pone.0158966
Genome-Wide Analysis of the bZIP Gene Family Identifies Two ABI5-Like bZIP Transcription Factors, BrABI5a and BrABI5b, as Positive Modulators of ABA Signalling in Chinese Cabbage
Abstract
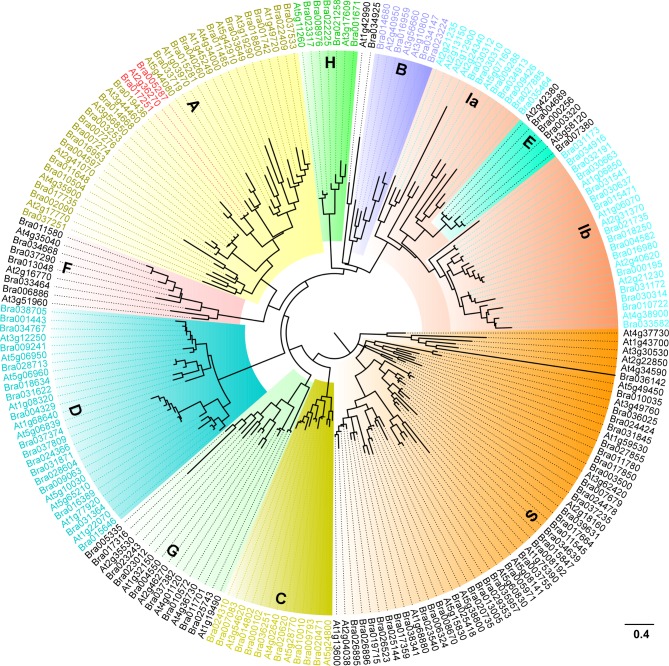
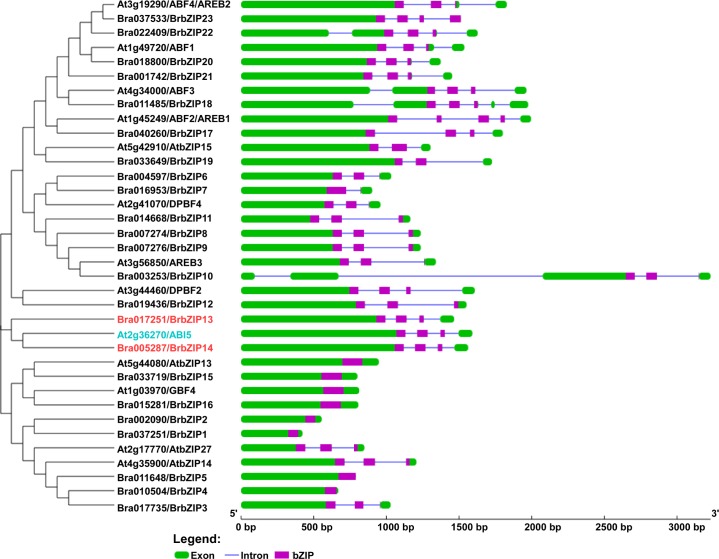
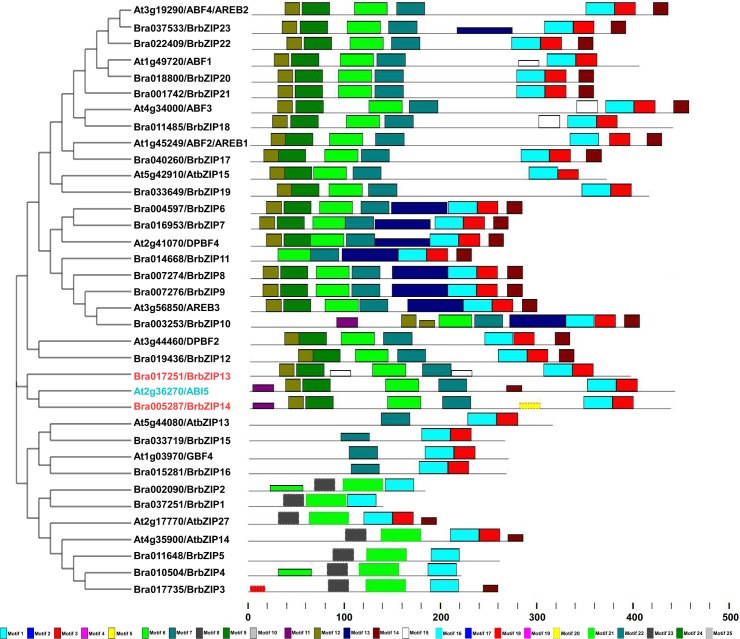
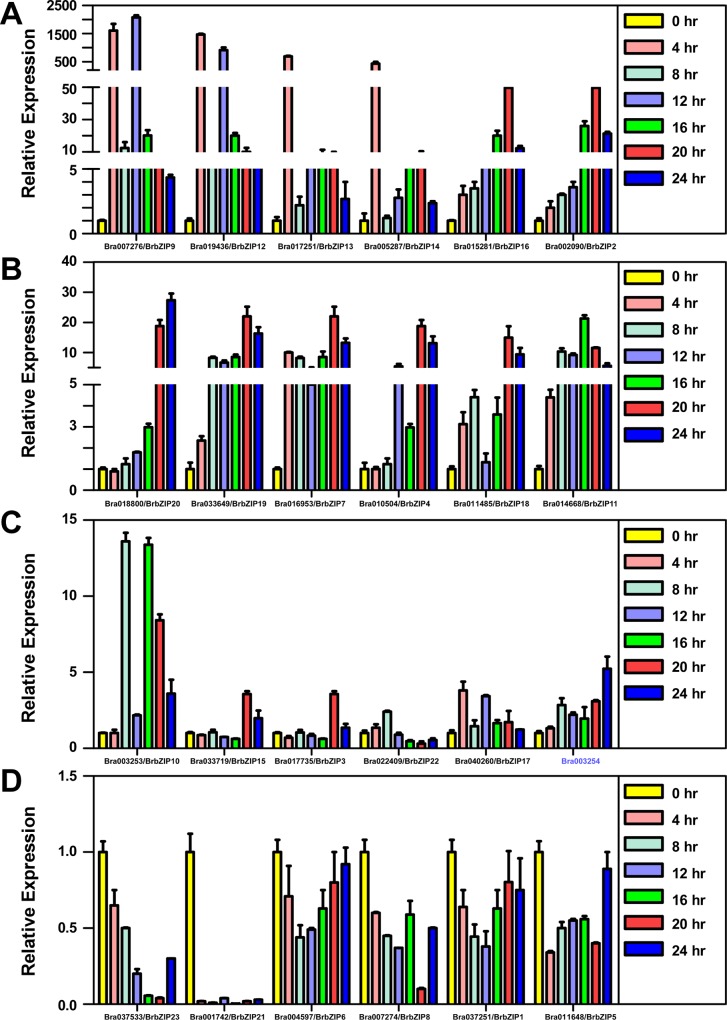
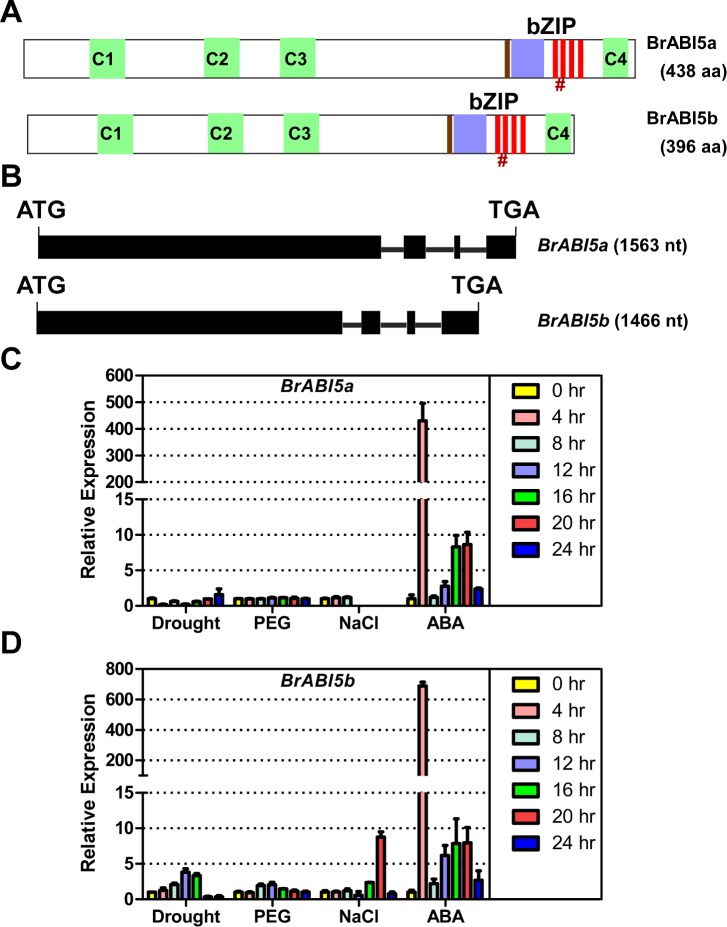
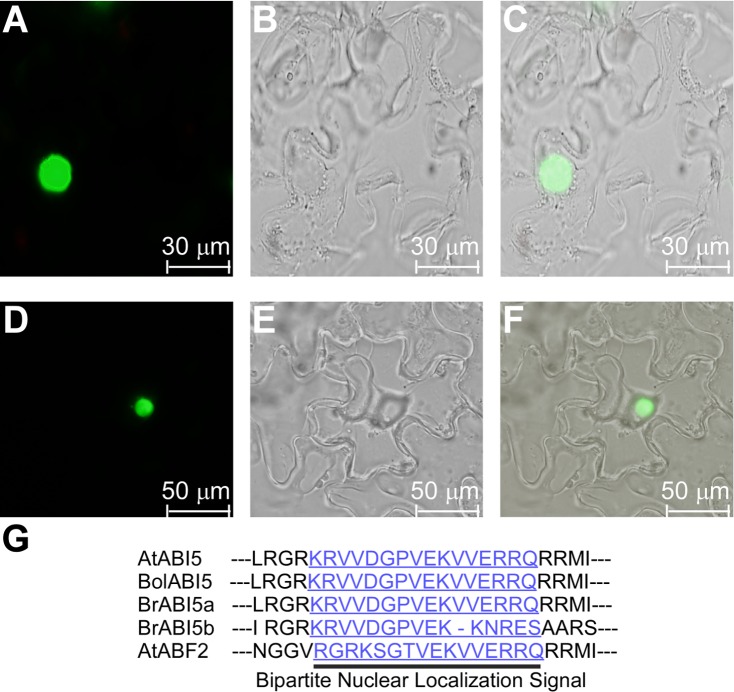
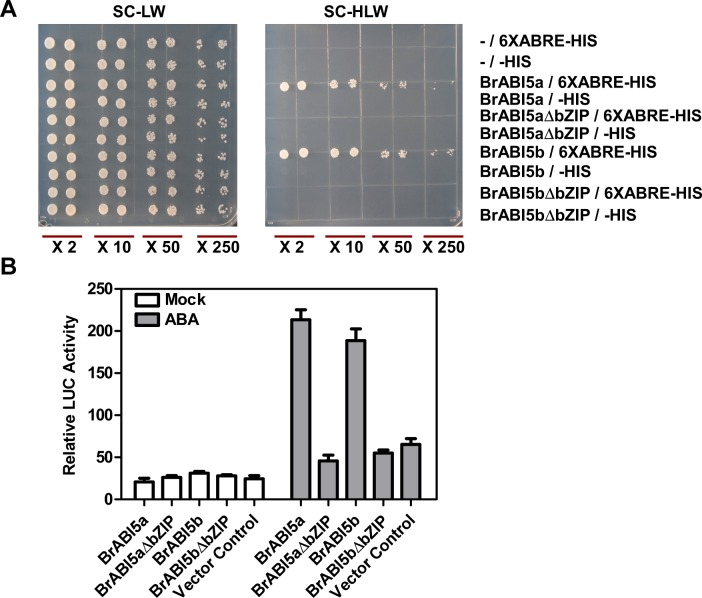
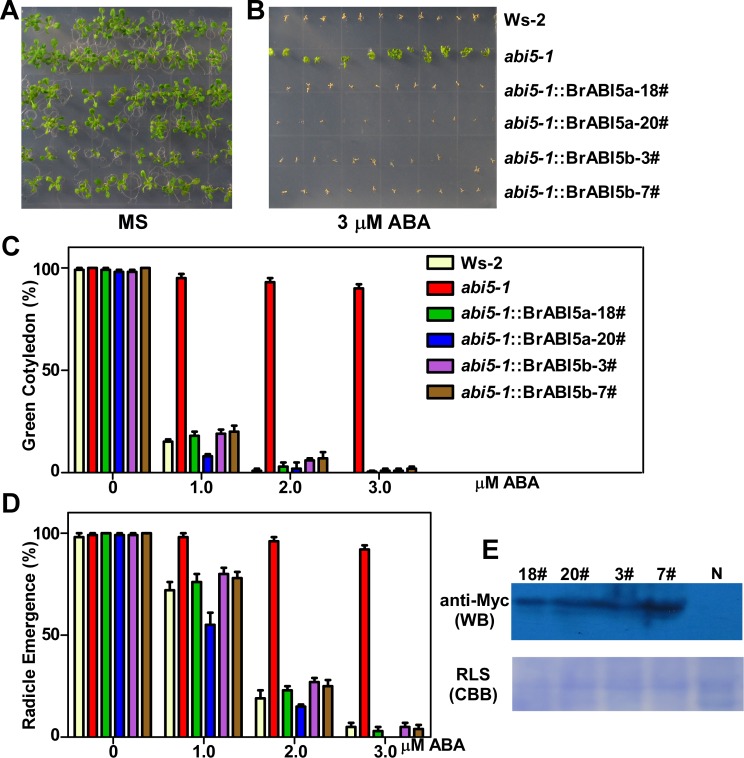
bZIP (basic leucine zipper) transcription factors coordinate plant growth and development and control responses to environmental stimuli. The genome of Chinese cabbage (Brassica rapa) encodes 136 putative bZIP transcription factors. The bZIP transcription factors in Brassica rapa (BrbZIP) are classified into 10 subfamilies. Phylogenetic relationship analysis reveals that subfamily A consists of 23 BrbZIPs. Two BrbZIPs within subfamily A, Bra005287 and Bra017251, display high similarity to ABI5 (ABA Insensitive 5). Expression of subfamily A BrbZIPs, like BrABI5a (Bra005287/BrbZIP14) and BrABI5b (Bra017251/BrbZIP13), are significantly induced by the plant hormone ABA. Subcellular localization assay reveal that both BrABI5a and BrABI5b have a nuclear localization. BrABI5a and BrABI5b could directly stimulate ABA Responsive Element-driven HIS (a HIS3 reporter gene, which confers His prototrophy) or LUC (LUCIFERASE) expression in yeast and Arabidopsis protoplast. Deletion of the bZIP motif abolished BrABI5a and BrABI5b transcriptional activity. The ABA insensitive phenotype of Arabidopsis abi5-1 is completely suppressed in transgenic lines expressing BrABI5a or BrABI5b. Overall, these results suggest that ABI5 orthologs, BrABI5a and BrABI5b, have key roles in ABA signalling in Chinese cabbage.
Conflict of interest statement
Figures
References
-
- Riechmann JL, Heard J, Martin G, Reuber L, Jiang C, Keddie J, et al. (2000) Arabidopsis transcription factors: genome-wide comparative analysis among eukaryotes. Science 290: 2105–2110. - PubMed
-
- Jakoby M, Weisshaar B, Droge-Laser W, Vicente-Carbajosa J, Tiedemann J, Kroj T, et al. (2002) bZIP transcription factors in Arabidopsis. Trends Plant Sci 7: 106–111. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
