

Local Epidermal Growth Factor Receptor Signaling Mediates the Systemic Pathogenic Effects of Staphylococcus aureus Toxic Shock Syndrome
- PMID: 27414801
- PMCID: PMC4944920
- DOI: 10.1371/journal.pone.0158969
Local Epidermal Growth Factor Receptor Signaling Mediates the Systemic Pathogenic Effects of Staphylococcus aureus Toxic Shock Syndrome
Abstract
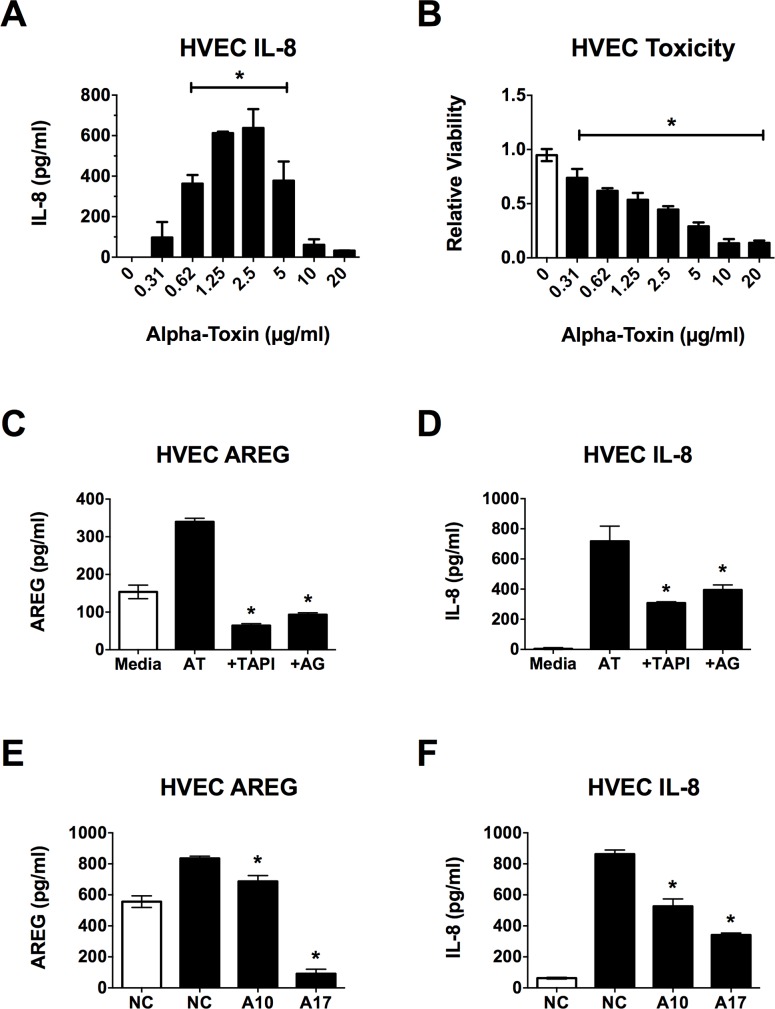
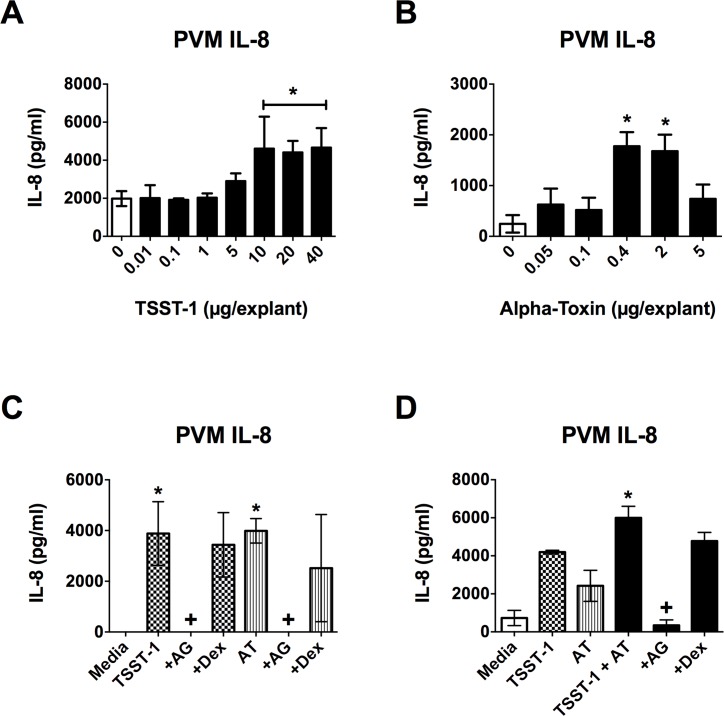
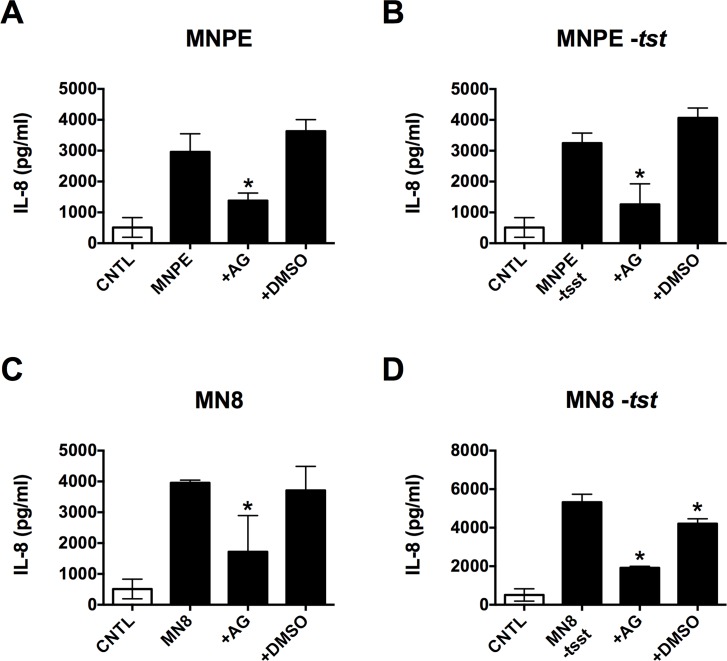
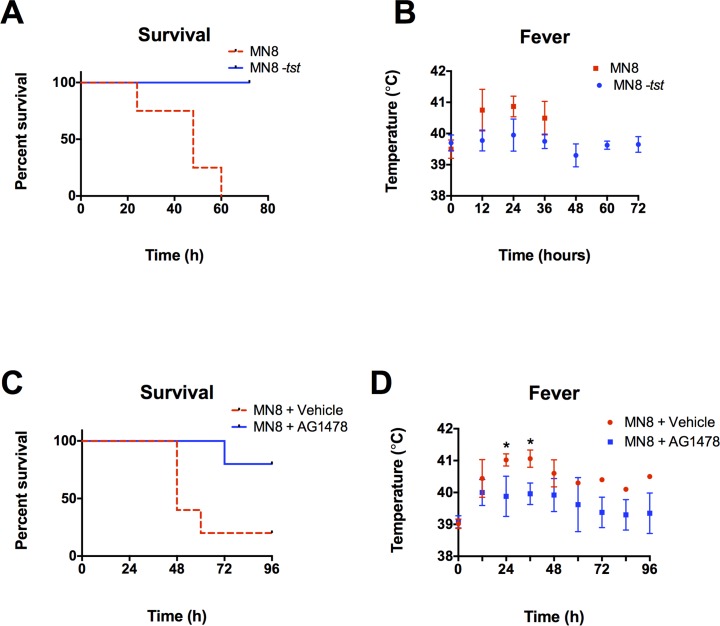
Secreted factors of Staphylococcus aureus can activate host signaling from the epidermal growth factor receptor (EGFR). The superantigen toxic shock syndrome toxin-1 (TSST-1) contributes to mucosal cytokine production through a disintegrin and metalloproteinase (ADAM)-mediated shedding of EGFR ligands and subsequent EGFR activation. The secreted hemolysin, α-toxin, can also induce EGFR signaling and directly interacts with ADAM10, a sheddase of EGFR ligands. The current work explores the role of EGFR signaling in menstrual toxic shock syndrome (mTSS), a disease mediated by TSST-1. The data presented show that TSST-1 and α-toxin induce ADAM- and EGFR-dependent cytokine production from human vaginal epithelial cells. TSST-1 and α-toxin also induce cytokine production from an ex vivo porcine vaginal mucosa (PVM) model. EGFR signaling is responsible for the majority of IL-8 production from PVM in response to secreted toxins and live S. aureus. Finally, data are presented demonstrating that inhibition of EGFR signaling with the EGFR-specific tyrosine kinase inhibitor AG1478 significantly increases survival in a rabbit model of mTSS. These data indicate that EGFR signaling is critical for progression of an S. aureus exotoxin-mediated disease and may represent an attractive host target for therapeutics.
Conflict of interest statement
Figures
References
-
- Huovila A-PJ, Turner AJ, Pelto-Huikko M, Kärkkäinen I, Ortiz RM. Shedding light on ADAM metalloproteinases. Trends Biochem Sci. 2005;30:413–422. - PubMed
-
- Pastore S, Mascia F, Mariani V, Girolomoni G. The epidermal growth factor receptor system in skin repair and inflammation. J Invest Dermatol. 2008;128:1365–1374. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
