

Endosome-ER Contacts Control Actin Nucleation and Retromer Function through VAP-Dependent Regulation of PI4P
- PMID: 27419871
- PMCID: PMC4963242
- DOI: 10.1016/j.cell.2016.06.037
Endosome-ER Contacts Control Actin Nucleation and Retromer Function through VAP-Dependent Regulation of PI4P
Abstract
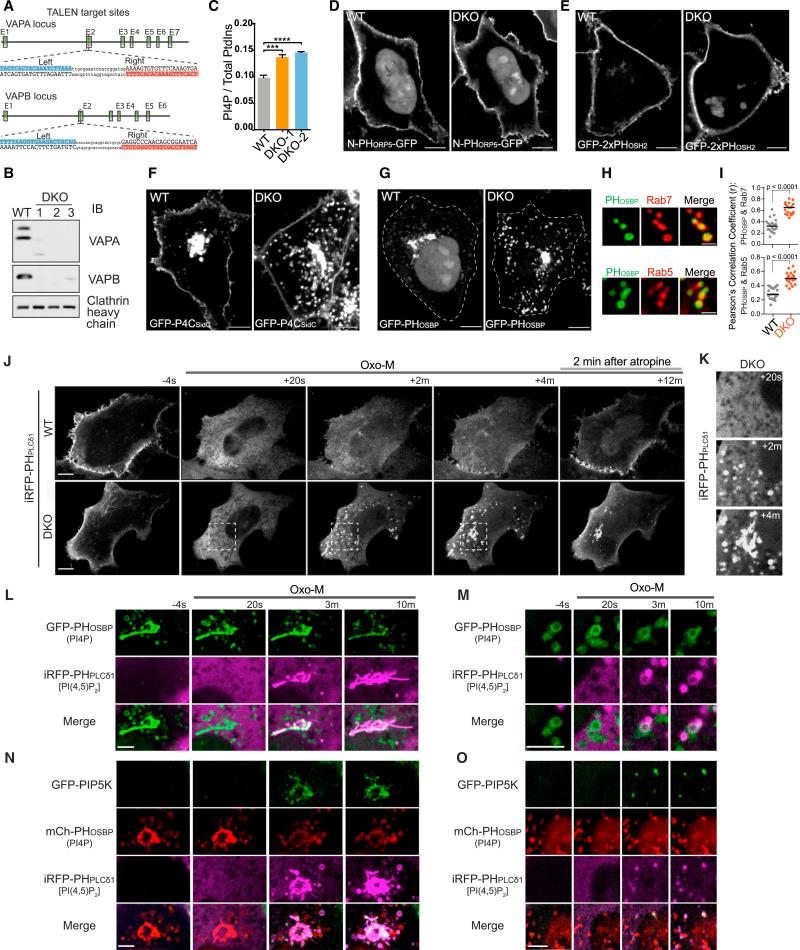
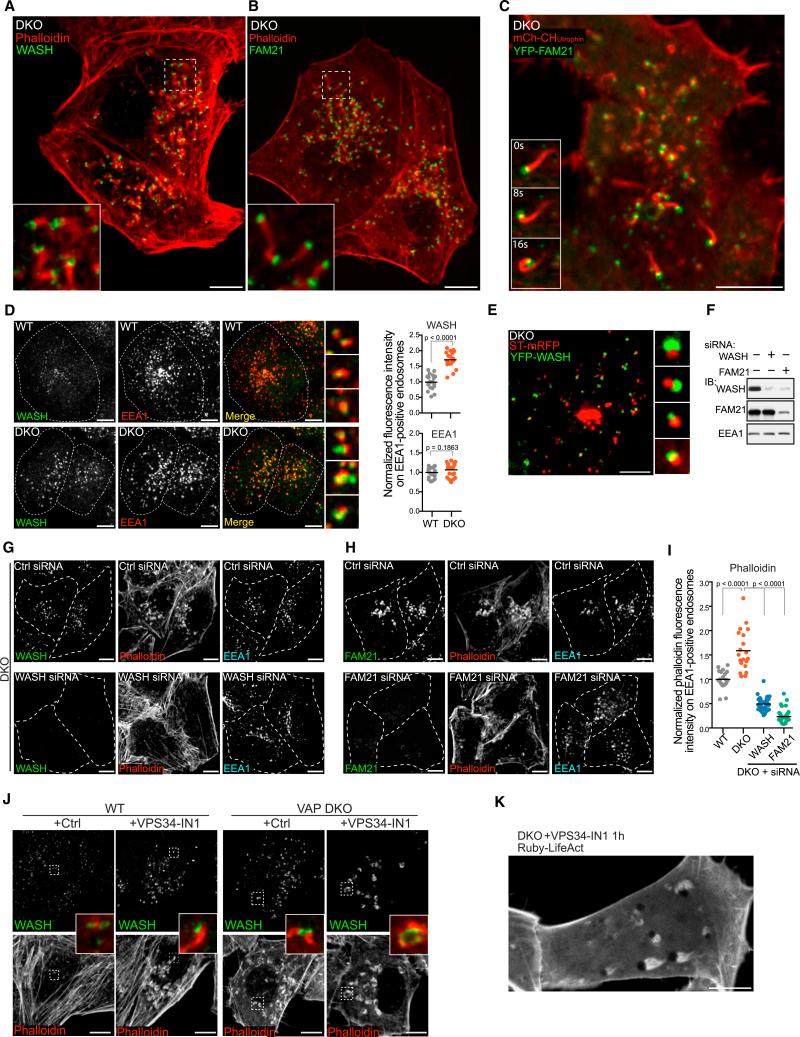
VAP (VAPA and VAPB) is an evolutionarily conserved endoplasmic reticulum (ER)-anchored protein that helps generate tethers between the ER and other membranes through which lipids are exchanged across adjacent bilayers. Here, we report that by regulating PI4P levels on endosomes, VAP affects WASH-dependent actin nucleation on these organelles and the function of the retromer, a protein coat responsible for endosome-to-Golgi traffic. VAP is recruited to retromer budding sites on endosomes via an interaction with the retromer SNX2 subunit. Cells lacking VAP accumulate high levels of PI4P, actin comets, and trans-Golgi proteins on endosomes. Such defects are mimicked by downregulation of OSBP, a VAP interactor and PI4P transporter that participates in VAP-dependent ER-endosomes tethers. These results reveal a role of PI4P in retromer-/WASH-dependent budding from endosomes. Collectively, our data show how the ER can control budding dynamics and association with the cytoskeleton of another membrane by direct contacts leading to bilayer lipid modifications.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures





References
-
- Alpy F, Rousseau A, Schwab Y, Legueux F, Stoll I, Wendling C, Spiegelhalter C, Kessler P, Mathelin C, Rio M-C, et al. STARD3 or STARD3NL and VAP form a novel molecular tether between late endosomes and the ER. J. Cell Sci. 2013;126:5500–5512. - PubMed
-
- Bago R, Malik N, Munson MJ, Prescott AR, Davies P, Sommer E, Shpiro N, Ward R, Cross D, Ganley IG, Alessi DR. Characterization of VPS34-IN1, a selective inhibitor of Vps34, reveals that the phosphatidylinositol 3-phosphate-binding SGK3 protein kinase is a downstream target of class III phosphoinositide 3-kinase. Biochem. J. 2014;463:413–427. - PMC - PubMed
-
- Balla A, Tuymetova G, Barshishat M, Geiszt M, Balla T. Characterization of type II phosphatidylinositol 4-kinase isoforms reveals association of the enzymes with endosomal vesicular compartments. J. Biol. Chem. 2002;277:20041–20050. - PubMed
-
- Bojjireddy N, Botyanszki J, Hammond G, Creech D, Peterson R, Kemp DC, Snead M, Brown R, Morrison A, Wilson S, et al. Pharmacological and genetic targeting of the PI4KA enzyme reveals its important role in maintaining plasma membrane phosphatidylinositol 4-phosphate and phosphatidylinositol 4,5-bisphosphate levels. J. Biol. Chem. 2014;289:6120–6132. - PMC - PubMed
-
- Bonifacino JS, Rojas R. Retrograde transport from endosomes to the trans-Golgi network. Nat. Rev. Mol. Cell Biol. 2006;7:568–579. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
