

Regulation of a polyamine transporter by the conserved 3' UTR-derived sRNA SorX confers resistance to singlet oxygen and organic hydroperoxides in Rhodobacter sphaeroides
- PMID: 27420112
- PMCID: PMC5056773
- DOI: 10.1080/15476286.2016.1212152
Regulation of a polyamine transporter by the conserved 3' UTR-derived sRNA SorX confers resistance to singlet oxygen and organic hydroperoxides in Rhodobacter sphaeroides
Abstract
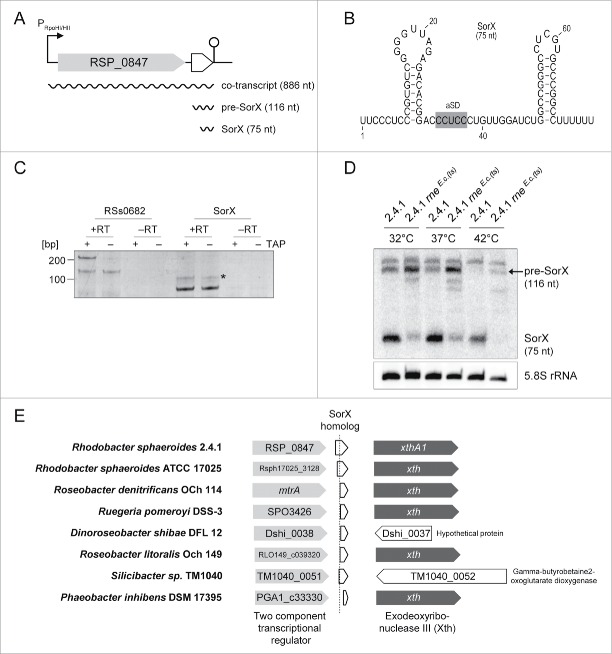
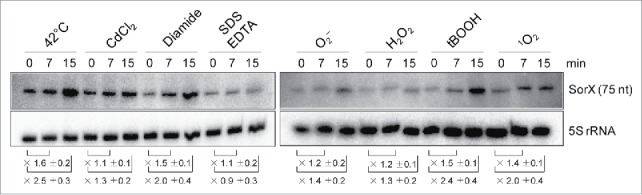
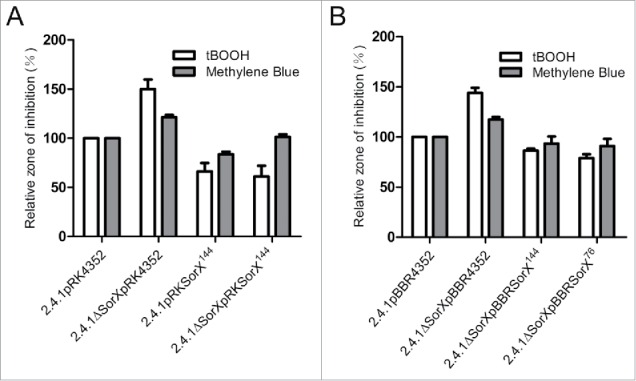
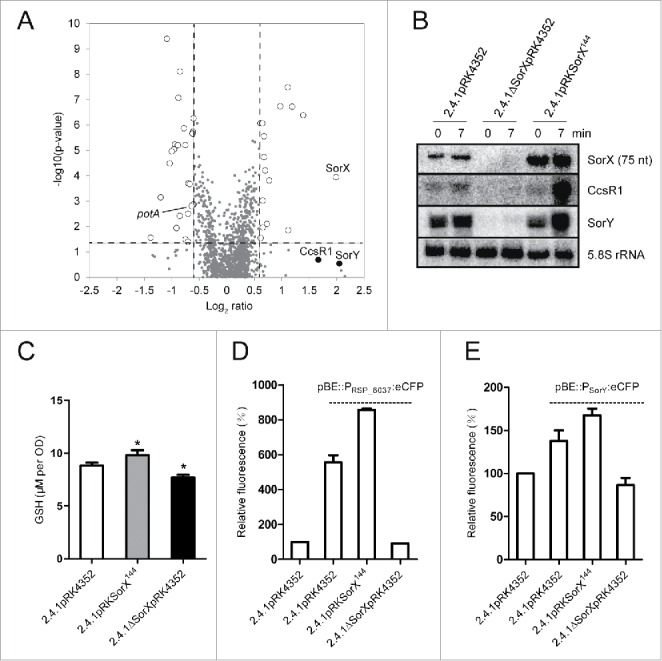
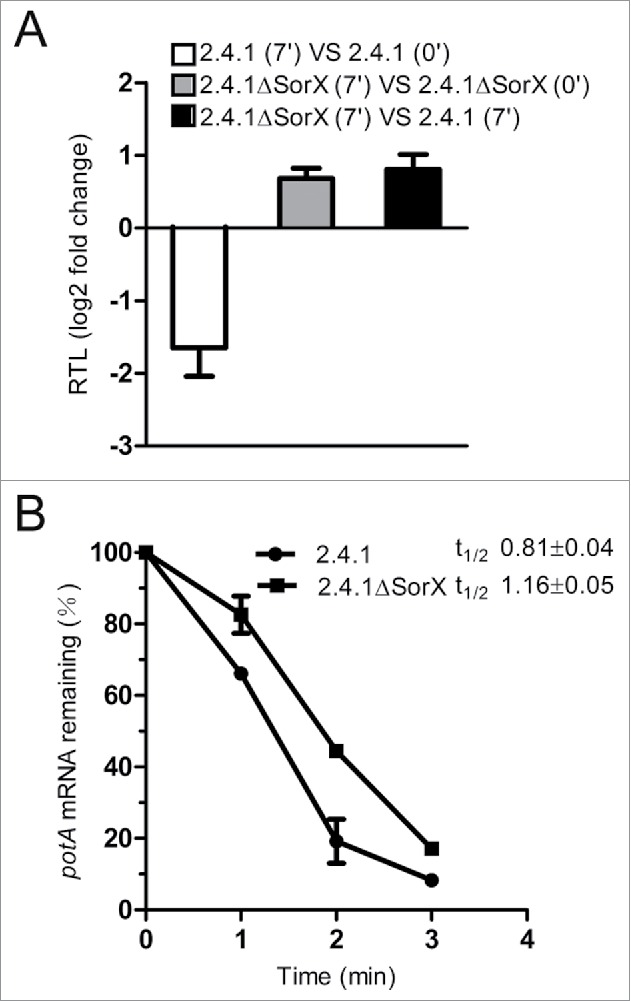
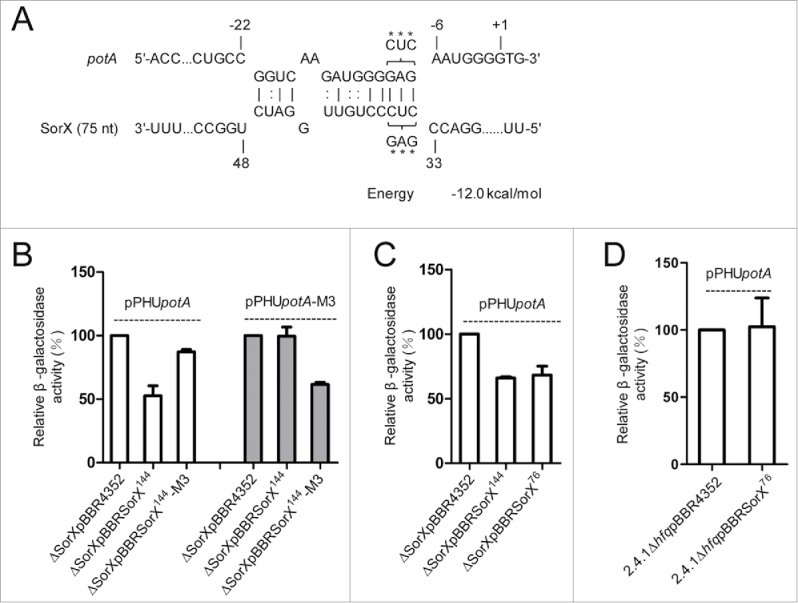
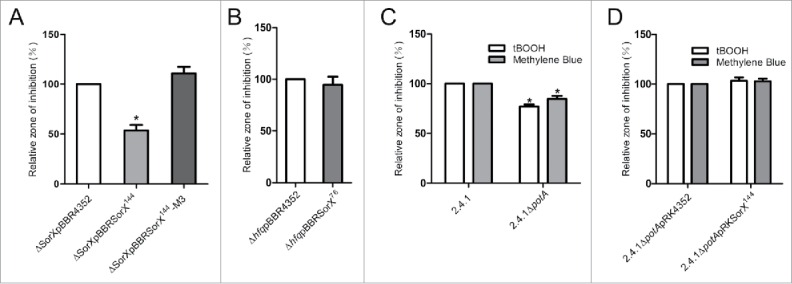
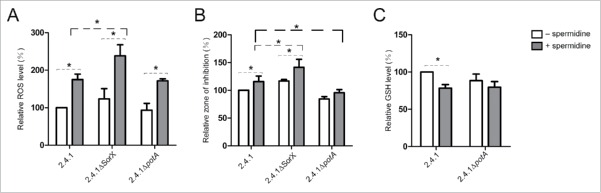
Singlet oxygen is generated by bacteriochlorophylls when light and oxygen are simultaneously present in Rhodobacter sphaeroides. Singlet oxygen triggers a specific response that is partly regulated by the alternative sigma factor RpoHI/HII. The sRNA RSs2461 has previously been identified as an RpoHI/HII-dependent sRNA and is derived from the 3' UTR of the mRNA for an OmpR-type transcriptional regulator. Similar to the RpoHI/HII-dependent CcsR and SorY sRNAs, RSs2461 affects the resistance of R. sphaeroides against singlet oxygen and was therefore renamed here SorX. Furthermore, SorX has a strong impact on resistance against organic hydroperoxides that usually occur as secondary damages downstream of singlet oxygen. The 75-nt SorX 3' fragment, which is generated by RNase E cleavage and highly conserved among related species, represents the functional entity. A target search identified potA mRNA, which encodes a subunit of a polyamine transporter, as a direct SorX target and stress resistance via SorX could be linked to potA. The PotABCD transporter is an uptake system for spermidine in E. coli. While spermidine is generally described as beneficial during oxidative stress, we observed significantly increased sensitivity of R. sphaeroides to organic hydroperoxides in the presence of spermidine. We therefore propose that the diminished import of spermidine, due to down-regulation of potA by SorX, counteracts oxidative stress. Together with results from other studies this underlines the importance of regulated transport to bacterial stress defense.
Keywords: 3′ UTR-derived; Hfq; Rhodobacter sphaeroides; organic hydroperoxide; polyamine transporter; potA; singlet oxygen; small RNA.
Figures
Similar articles
-
The sRNA SorY confers resistance during photooxidative stress by affecting a metabolite transporter in Rhodobacter sphaeroides.RNA Biol. 2015;12(5):569-77. doi: 10.1080/15476286.2015.1031948. RNA Biol. 2015. PMID: 25833751 Free PMC article.
-
A response regulator of the OmpR family is part of the regulatory network controlling the oxidative stress response of Rhodobacter sphaeroides.Environ Microbiol Rep. 2019 Apr;11(2):118-128. doi: 10.1111/1758-2229.12718. Epub 2018 Dec 3. Environ Microbiol Rep. 2019. PMID: 30451391
-
Singlet oxygen stress in microorganisms.Adv Microb Physiol. 2011;58:141-73. doi: 10.1016/B978-0-12-381043-4.00004-0. Adv Microb Physiol. 2011. PMID: 21722793 Review.
-
A cluster of four homologous small RNAs modulates C1 metabolism and the pyruvate dehydrogenase complex in Rhodobacter sphaeroides under various stress conditions.J Bacteriol. 2015 May;197(10):1839-52. doi: 10.1128/JB.02475-14. Epub 2015 Mar 16. J Bacteriol. 2015. PMID: 25777678 Free PMC article.
-
Shedding light on a Group IV (ECF11) alternative σ factor.Mol Microbiol. 2019 Aug;112(2):374-384. doi: 10.1111/mmi.14280. Epub 2019 May 31. Mol Microbiol. 2019. PMID: 31111523 Free PMC article. Review.
Cited by
-
Maturation of UTR-Derived sRNAs Is Modulated during Adaptation to Different Growth Conditions.Int J Mol Sci. 2021 Nov 12;22(22):12260. doi: 10.3390/ijms222212260. Int J Mol Sci. 2021. PMID: 34830143 Free PMC article.
-
The protein regulator ArgR and the sRNA derived from the 3'-UTR region of its gene, ArgX, both regulate the arginine deiminase pathway in Lactococcus lactis.PLoS One. 2019 Jun 20;14(6):e0218508. doi: 10.1371/journal.pone.0218508. eCollection 2019. PLoS One. 2019. PMID: 31220124 Free PMC article.
-
Ribonuclease E strongly impacts bacterial adaptation to different growth conditions.RNA Biol. 2023 Jan;20(1):120-135. doi: 10.1080/15476286.2023.2195733. RNA Biol. 2023. PMID: 36988476 Free PMC article.
-
Regulatory 3' Untranslated Regions of Bacterial mRNAs.Front Microbiol. 2017 Jul 10;8:1276. doi: 10.3389/fmicb.2017.01276. eCollection 2017. Front Microbiol. 2017. PMID: 28740488 Free PMC article. Review.
-
Adaptation to Photooxidative Stress: Common and Special Strategies of the Alphaproteobacteria Rhodobacter sphaeroides and Rhodobacter capsulatus.Microorganisms. 2020 Feb 19;8(2):283. doi: 10.3390/microorganisms8020283. Microorganisms. 2020. PMID: 32093084 Free PMC article.
References
-
- Bauer C, Elsen S, Swem LR, Swem DL, Masuda S. Redox and light regulation of gene expression in photosynthetic prokaryotes. Philos Trans R Soc Lond B Biol Sci 2003; 358:147-53; PMID:12594923; http://dx.doi.org/10.1098/rstb.2002.1189 - DOI - PMC - PubMed
-
- Glaeser J, Klug G. Photo-oxidative stress in Rhodobacter sphaeroides: protective role of carotenoids and expression of selected genes. Microbiol 2005; 151:1927-38; PMID:15942000; http://dx.doi.org/1872302710.1099/mic.0.27789-0 - DOI - PubMed
-
- Dufour YS, Landick R, Donohue TJ. Organization and evolution of the biological response to singlet oxygen stress. J Mol Biol 2008; 383:713-30; PMID:18723027; http://dx.doi.org/10.1016/j.jmb.2008.08.017 - DOI - PMC - PubMed
-
- Glaeser J, Nuss AM, Berghoff BA, Klug G. Singlet oxygen stress in microorganisms. Adv Microb Physiol 2011; 58:141-73; PMID:21722793; http://dx.doi.org/10.1016/B978-0-12-381043-4.00004-0 - DOI - PubMed
-
- Glaeser J, Zobawa M, Lottspeich F, Klug G. Protein synthesis patterns reveal a complex regulatory response to singlet oxygen in Rhodobacter. J Proteome Res 2007; 6:2460-71; PMID:17536848; http://dx.doi.org/10.1021/pr060624p - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials