

OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2 O2 signalling in rice
- PMID: 27420922
- PMCID: PMC5258865
- DOI: 10.1111/pbi.12601
OsASR5 enhances drought tolerance through a stomatal closure pathway associated with ABA and H2 O2 signalling in rice
Abstract
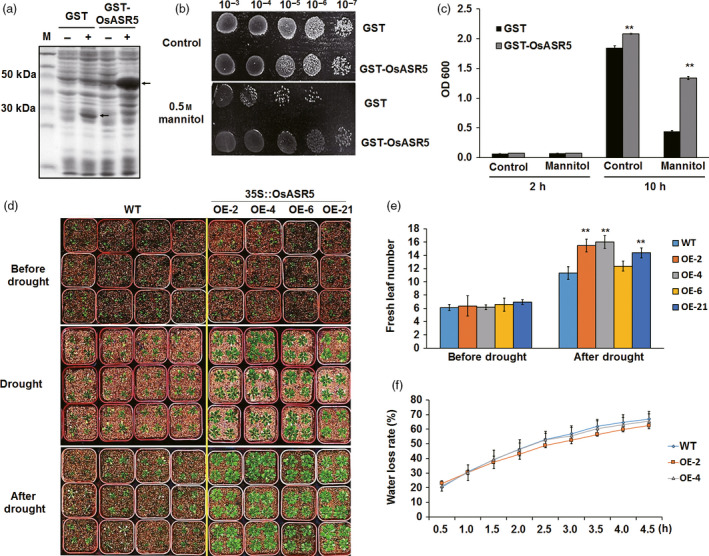
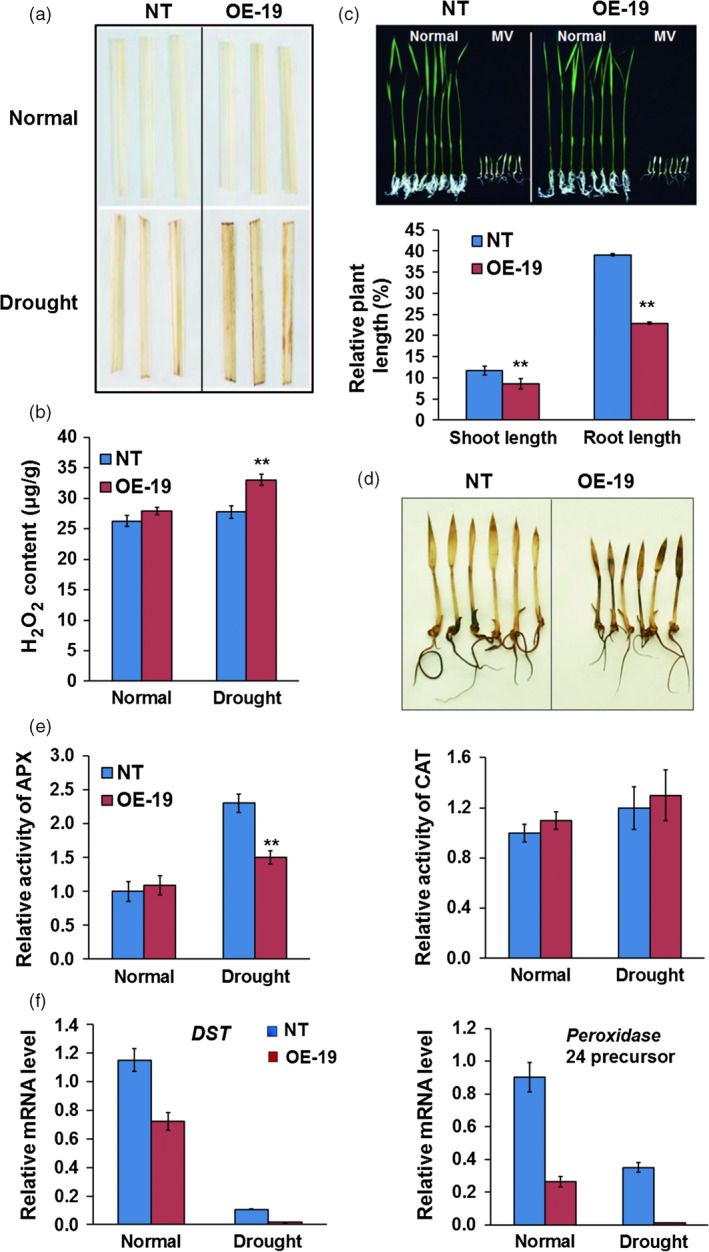
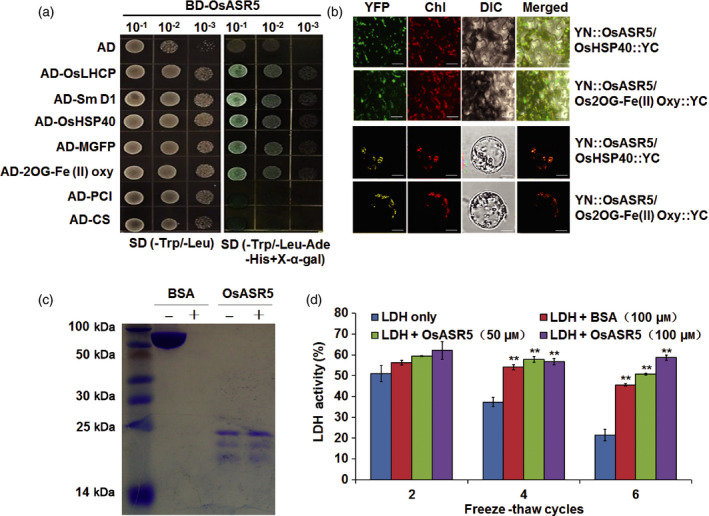
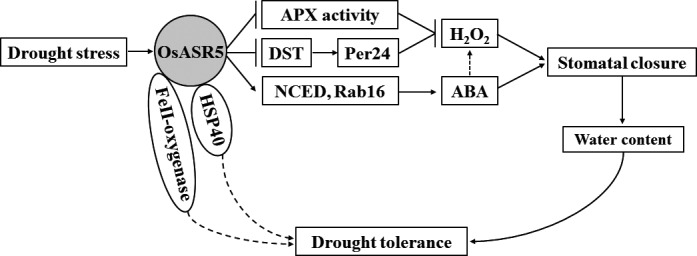
Drought is one of the major abiotic stresses that directly implicate plant growth and crop productivity. Although many genes in response to drought stress have been identified, genetic improvement to drought resistance especially in food crops is showing relatively slow progress worldwide. Here, we reported the isolation of abscisic acid, stress and ripening (ASR) genes from upland rice variety, IRAT109 (Oryza sativa L. ssp. japonica), and demonstrated that overexpression of OsASR5 enhanced osmotic tolerance in Escherichia coli and drought tolerance in Arabidopsis and rice by regulating leaf water status under drought stress conditions. Moreover, overexpression of OsASR5 in rice increased endogenous ABA level and showed hypersensitive to exogenous ABA treatment at both germination and postgermination stages. The production of H2 O2 , a second messenger for the induction of stomatal closure in response to ABA, was activated in overexpression plants under drought stress conditions, consequently, increased stomatal closure and decreased stomatal conductance. In contrast, the loss-of-function mutant, osasr5, showed sensitivity to drought stress with lower relative water content under drought stress conditions. Further studies demonstrated that OsASR5 functioned as chaperone-like protein and interacted with stress-related HSP40 and 2OG-Fe (II) oxygenase domain containing proteins in yeast and plants. Taken together, we suggest that OsASR5 plays multiple roles in response to drought stress by regulating ABA biosynthesis, promoting stomatal closure, as well as acting as chaperone-like protein that possibly prevents drought stress-related proteins from inactivation.
Keywords: ABA; Oryza sativa; OsASR5; Drought; stomata; water content.
© 2016 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures






References
-
- Cho, E.K. and Hong, C.B. (2006) Over‐expression of tobacco NtHSP70‐1 contributes to drought‐stress tolerance in plants. Plant Cell Rep. 25, 349–358. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
