

Processive DNA Unwinding by RecBCD Helicase in the Absence of Canonical Motor Translocation
- PMID: 27422010
- PMCID: PMC4976051
- DOI: 10.1016/j.jmb.2016.07.002
Processive DNA Unwinding by RecBCD Helicase in the Absence of Canonical Motor Translocation
Abstract
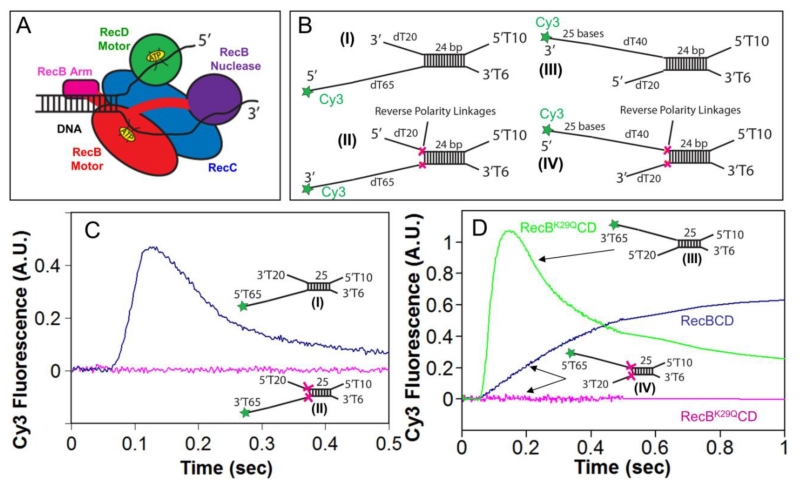
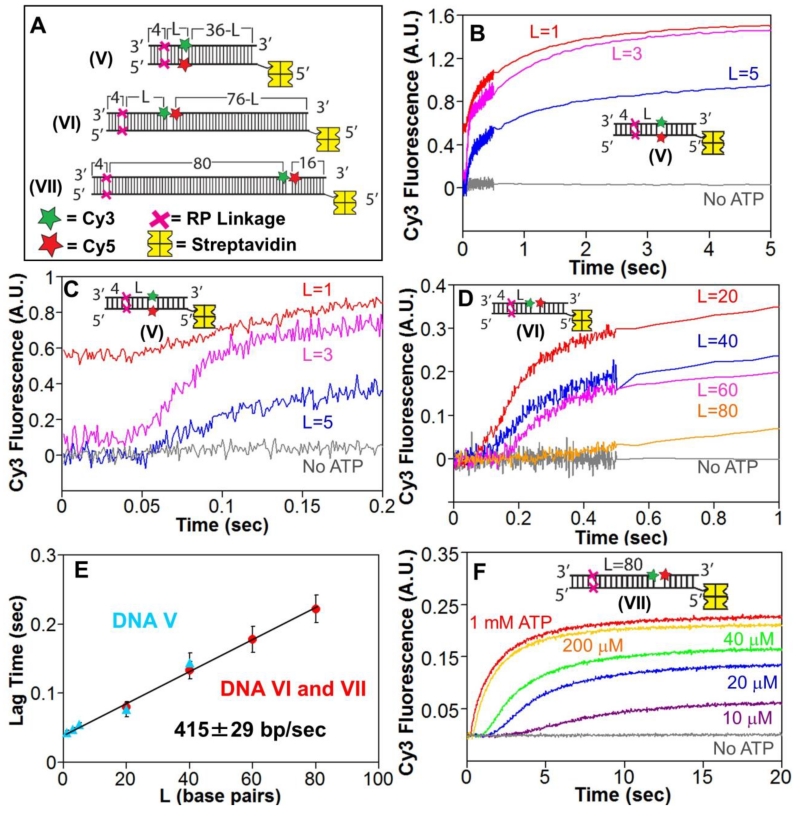
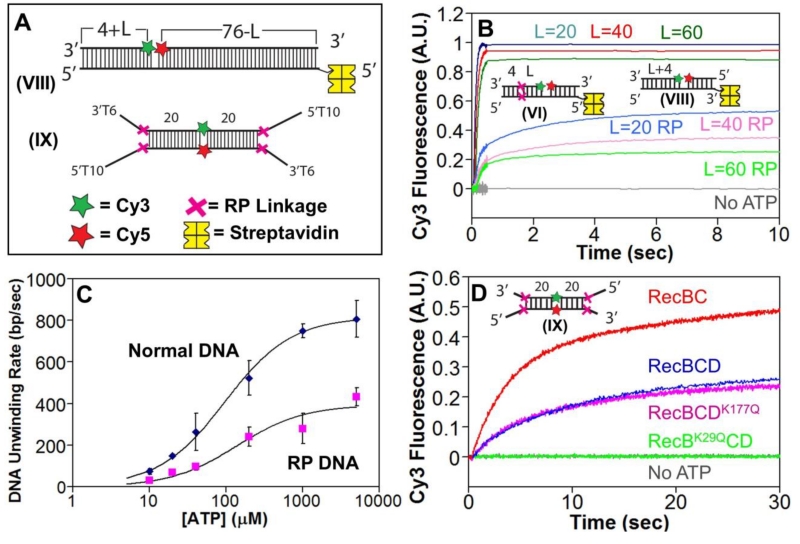
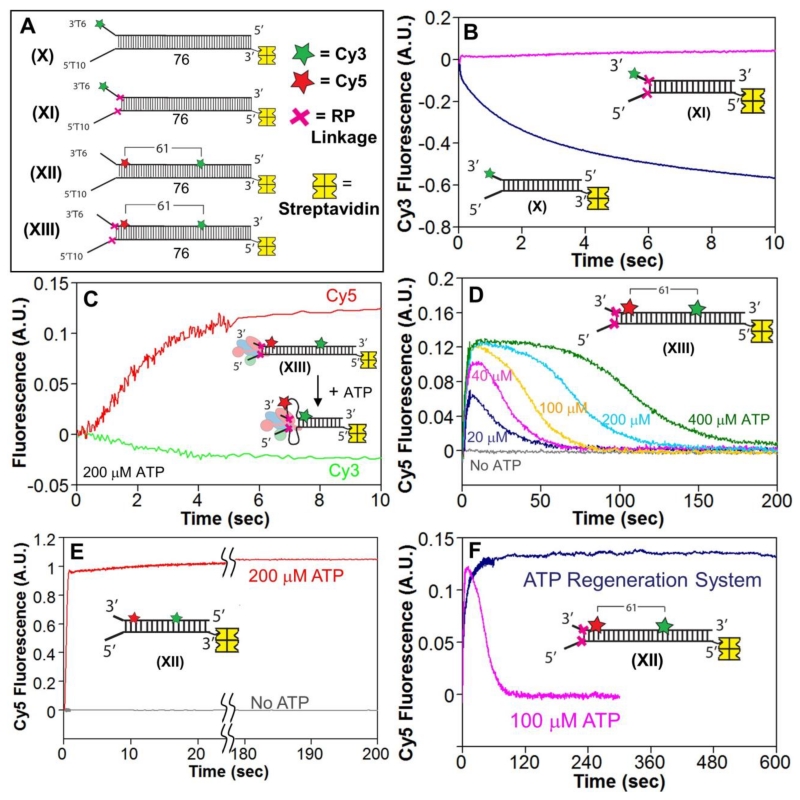
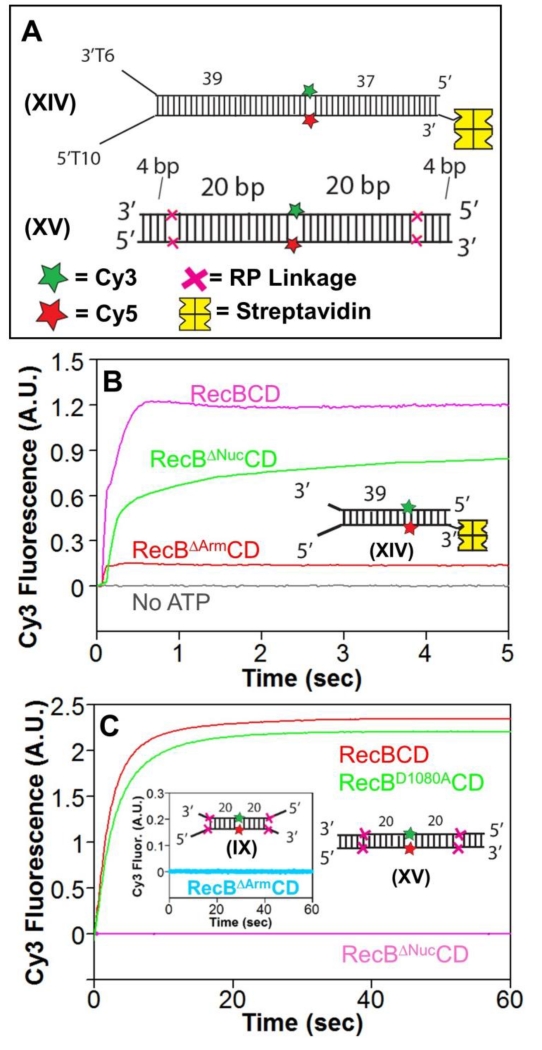
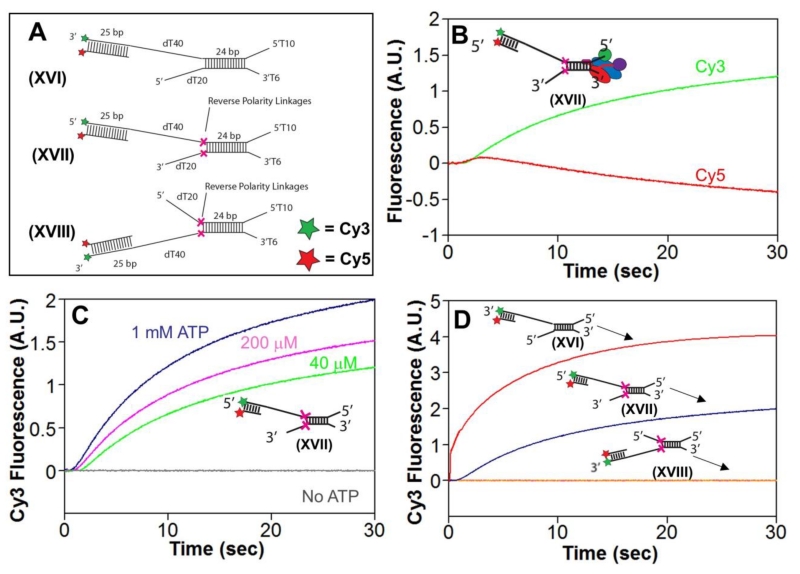
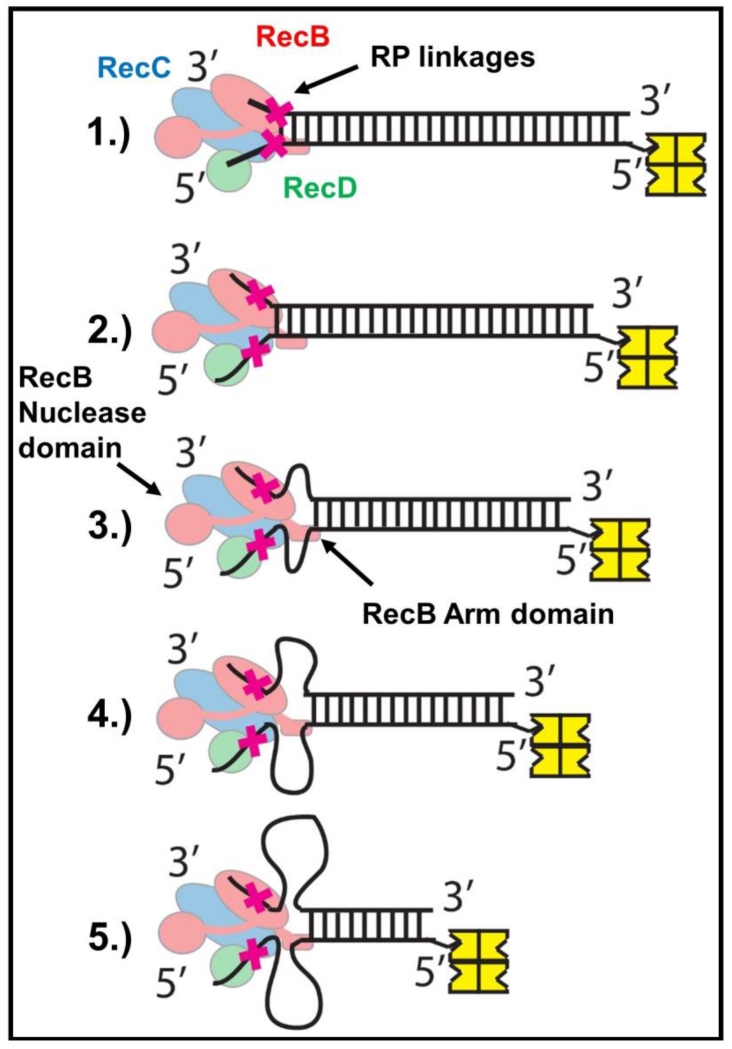
Escherichia coli RecBCD is a DNA helicase/nuclease that functions in double-stranded DNA break repair. RecBCD possesses two motors (RecB, a 3' to 5' translocase, and RecD, a 5' to 3' translocase). Current DNA unwinding models propose that motor translocation is tightly coupled to base pair melting. However, some biochemical evidence suggests that DNA melting of multiple base pairs may occur separately from single-stranded DNA translocation. To test this hypothesis, we designed DNA substrates containing reverse backbone polarity linkages that prevent ssDNA translocation of the canonical RecB and RecD motors. Surprisingly, we find that RecBCD can processively unwind DNA for at least 80bp beyond the reverse polarity linkages. This ability requires an ATPase active RecB motor, the RecB "arm" domain, and also the RecB nuclease domain, but not its nuclease activity. These results indicate that RecBCD can unwind duplex DNA processively in the absence of ssDNA translocation by the canonical motors and that the nuclease domain regulates the helicase activity of RecBCD.
Keywords: SF1 helicase; allostery; fluorescence; recombination.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
How Does a Helicase Unwind DNA? Insights from RecBCD Helicase.Bioessays. 2018 Jun;40(6):e1800009. doi: 10.1002/bies.201800009. Epub 2018 Mar 30. Bioessays. 2018. PMID: 29603305 Free PMC article. Review.
-
E. coli RecB Nuclease Domain Regulates RecBCD Helicase Activity but not Single Stranded DNA Translocase Activity.J Mol Biol. 2024 Jan 15;436(2):168381. doi: 10.1016/j.jmb.2023.168381. Epub 2023 Dec 9. J Mol Biol. 2024. PMID: 38081382 Free PMC article.
-
E. coli RecBCD Nuclease Domain Regulates Helicase Activity but not Single Stranded DNA Translocase Activity.bioRxiv [Preprint]. 2023 Oct 17:2023.10.13.561901. doi: 10.1101/2023.10.13.561901. bioRxiv. 2023. Update in: J Mol Biol. 2024 Jan 15;436(2):168381. doi: 10.1016/j.jmb.2023.168381. PMID: 37905078 Free PMC article. Updated. Preprint.
-
Heterogeneity in E. coli RecBCD Helicase-DNA Binding and Base Pair Melting.J Mol Biol. 2021 Sep 3;433(18):167147. doi: 10.1016/j.jmb.2021.167147. Epub 2021 Jul 9. J Mol Biol. 2021. PMID: 34246654 Free PMC article.
-
Escherichia coli DNA helicases: mechanisms of DNA unwinding.Mol Microbiol. 1992 Jan;6(1):5-14. doi: 10.1111/j.1365-2958.1992.tb00831.x. Mol Microbiol. 1992. PMID: 1310794 Review.
Cited by
-
Conformational selection and induced fit as a useful framework for molecular motor mechanisms.Biophys Chem. 2017 Apr;223:11-16. doi: 10.1016/j.bpc.2017.01.004. Epub 2017 Feb 3. Biophys Chem. 2017. PMID: 28187350 Free PMC article. Review.
-
How Does a Helicase Unwind DNA? Insights from RecBCD Helicase.Bioessays. 2018 Jun;40(6):e1800009. doi: 10.1002/bies.201800009. Epub 2018 Mar 30. Bioessays. 2018. PMID: 29603305 Free PMC article. Review.
-
Probing E. coli SSB protein-DNA topology by reversing DNA backbone polarity.Biophys J. 2021 Apr 20;120(8):1522-1533. doi: 10.1016/j.bpj.2021.02.025. Epub 2021 Feb 23. Biophys J. 2021. PMID: 33636169 Free PMC article.
-
Synergy between RecBCD subunits is essential for efficient DNA unwinding.Elife. 2019 Jan 2;8:e40836. doi: 10.7554/eLife.40836. Elife. 2019. PMID: 30601118 Free PMC article.
-
Communication between DNA and nucleotide binding sites facilitates stepping by the RecBCD helicase.Nucleic Acids Res. 2024 Apr 24;52(7):3911-3923. doi: 10.1093/nar/gkae108. Nucleic Acids Res. 2024. PMID: 38364872 Free PMC article.
References
-
- Singleton MR, Dillingham MS, Gaudier M, Kowalczykowski SC, Wigley DB. Crystal structure of RecBCD enzyme reveals a machine for processing DNA breaks. Nature. 2004;432:187–93. - PubMed
-
- Chen HW, Ruan B, Yu M, Wang J, Julin DA. The RecD subunit of the RecBCD enzyme from Escherichia coli is a single-stranded DNA-dependent ATPase. J Biol Chem. 1997;272:10072–9. - PubMed
-
- Taylor AF, Smith GR. RecBCD enzyme is a DNA helicase with fast and slow motors of opposite polarity. Nature. 2003;423:889–93. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
