

Automated Structure- and Sequence-Based Design of Proteins for High Bacterial Expression and Stability
- PMID: 27425410
- PMCID: PMC4961223
- DOI: 10.1016/j.molcel.2016.06.012
Automated Structure- and Sequence-Based Design of Proteins for High Bacterial Expression and Stability
Erratum in
-
Automated Structure- and Sequence-Based Design of Proteins for High Bacterial Expression and Stability.Mol Cell. 2018 Apr 19;70(2):380. doi: 10.1016/j.molcel.2018.03.035. Mol Cell. 2018. PMID: 29677494 Free PMC article. No abstract available.
Abstract
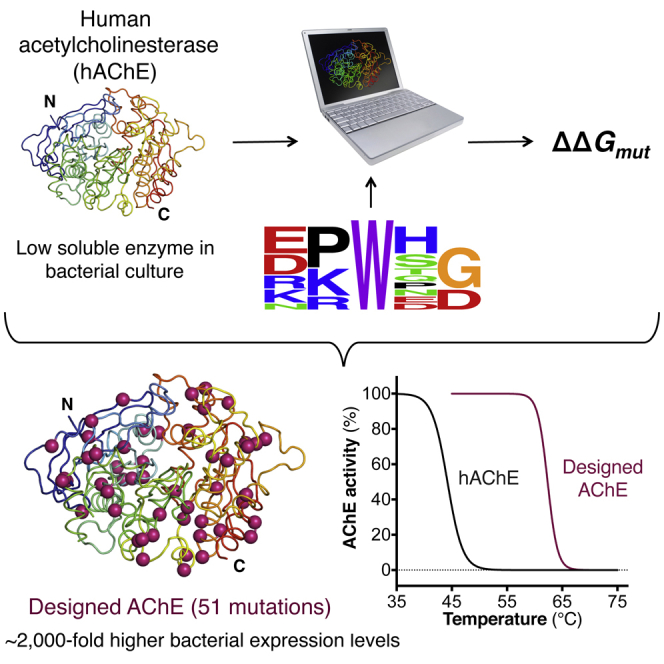
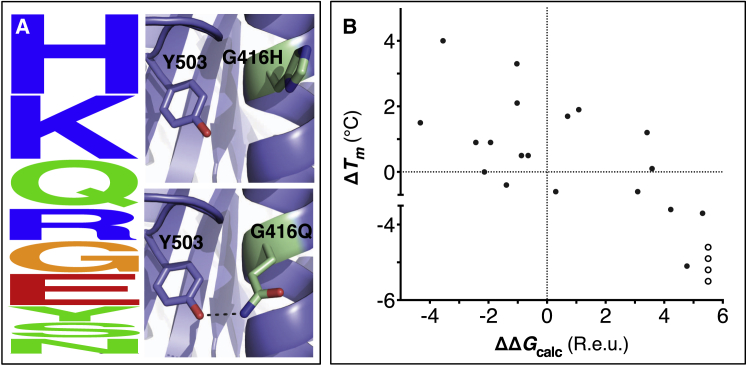
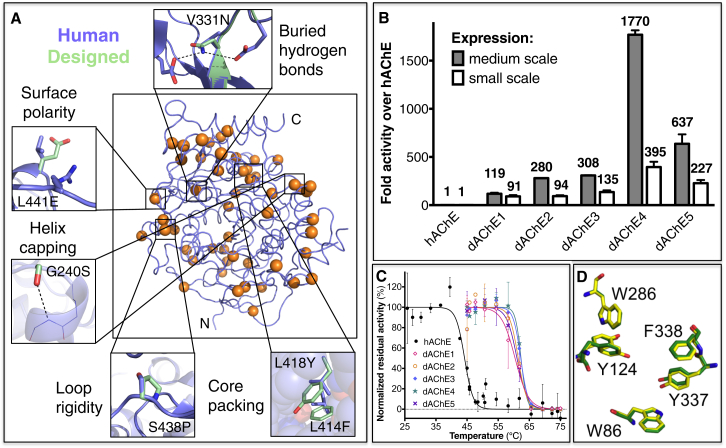
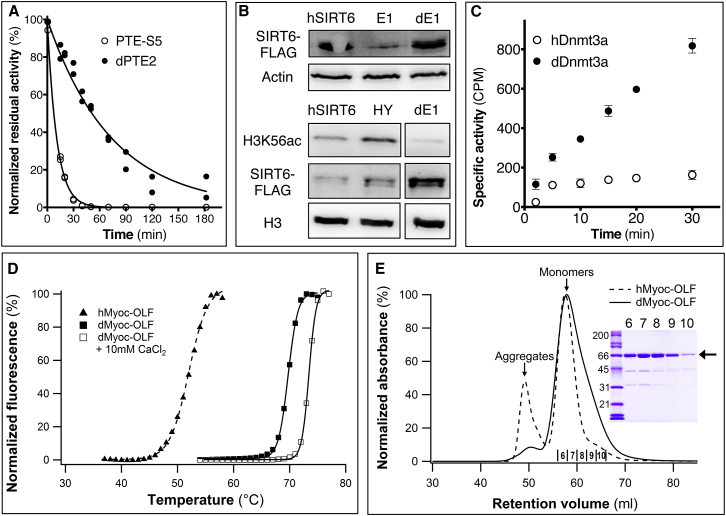
Upon heterologous overexpression, many proteins misfold or aggregate, thus resulting in low functional yields. Human acetylcholinesterase (hAChE), an enzyme mediating synaptic transmission, is a typical case of a human protein that necessitates mammalian systems to obtain functional expression. We developed a computational strategy and designed an AChE variant bearing 51 mutations that improved core packing, surface polarity, and backbone rigidity. This variant expressed at ∼2,000-fold higher levels in E. coli compared to wild-type hAChE and exhibited 20°C higher thermostability with no change in enzymatic properties or in the active-site configuration as determined by crystallography. To demonstrate broad utility, we similarly designed four other human and bacterial proteins. Testing at most three designs per protein, we obtained enhanced stability and/or higher yields of soluble and active protein in E. coli. Our algorithm requires only a 3D structure and several dozen sequences of naturally occurring homologs, and is available at http://pross.weizmann.ac.il.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Altschul S.F., Gish W., Miller W., Myers E.W., Lipman D.J. Basic local alignment search tool. J. Mol. Biol. 1990;215:403–410. - PubMed
-
- Benning M.M., Shim H., Raushel F.M., Holden H.M. High resolution X-ray structures of different metal-substituted forms of phosphotriesterase from Pseudomonas diminuta. Biochemistry. 2001;40:2712–2722. - PubMed
-
- Bigley A.N., Mabanglo M.F., Harvey S.P., Raushel F.M. Variants of phosphotriesterase for the enhanced detoxification of the chemical warfare agent VR. Biochemistry. 2015;54:5502–5512. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
