

Systems Analysis of the Dynamic Inflammatory Response to Tissue Damage Reveals Spatiotemporal Properties of the Wound Attractant Gradient
- PMID: 27426513
- PMCID: PMC4985561
- DOI: 10.1016/j.cub.2016.06.012
Systems Analysis of the Dynamic Inflammatory Response to Tissue Damage Reveals Spatiotemporal Properties of the Wound Attractant Gradient
Abstract
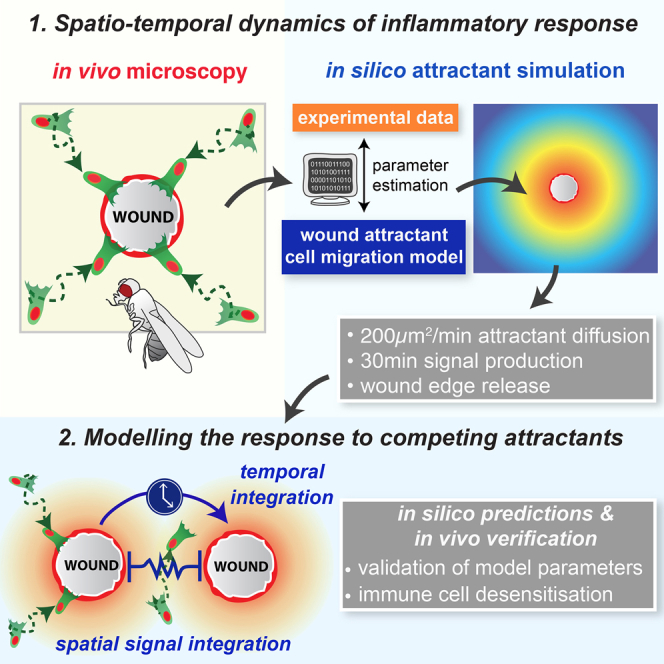
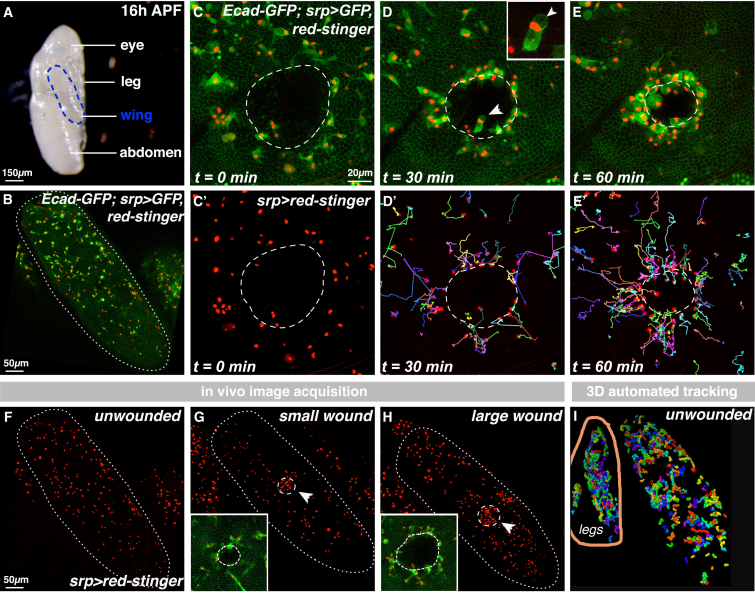
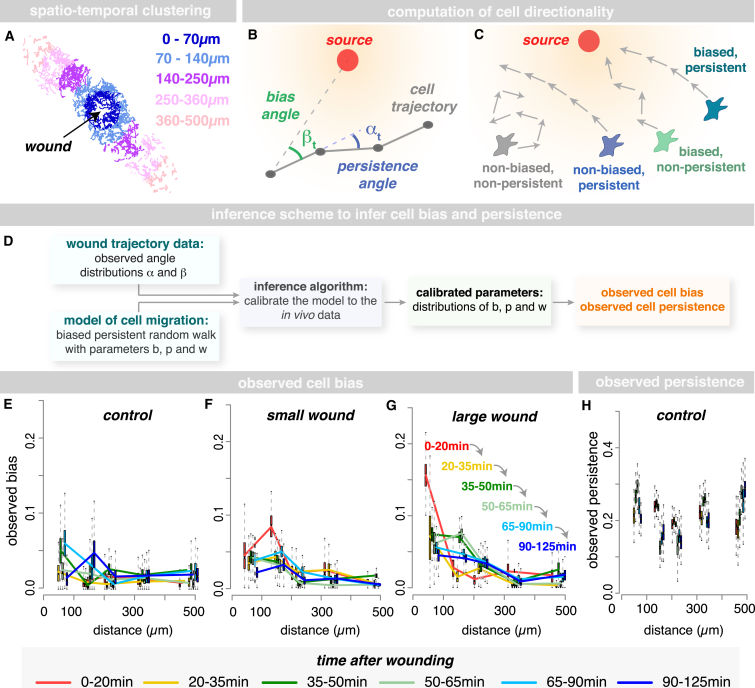
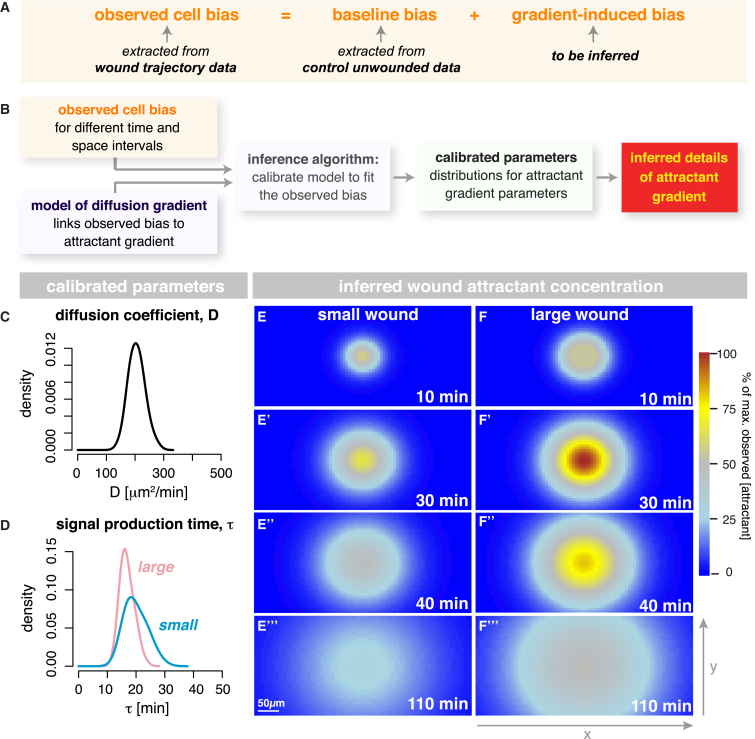
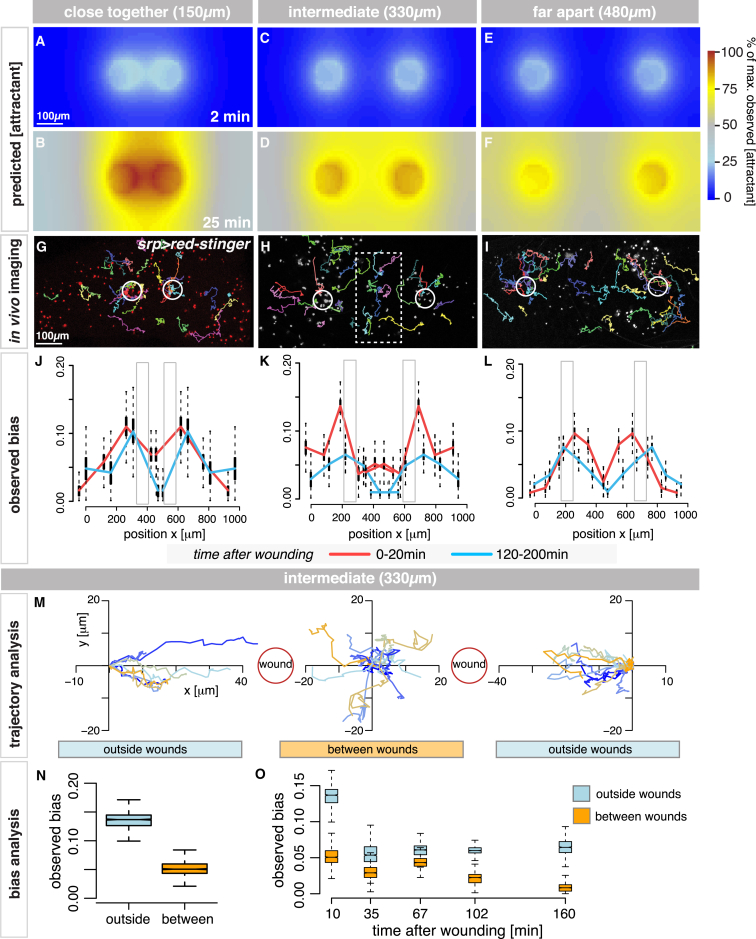
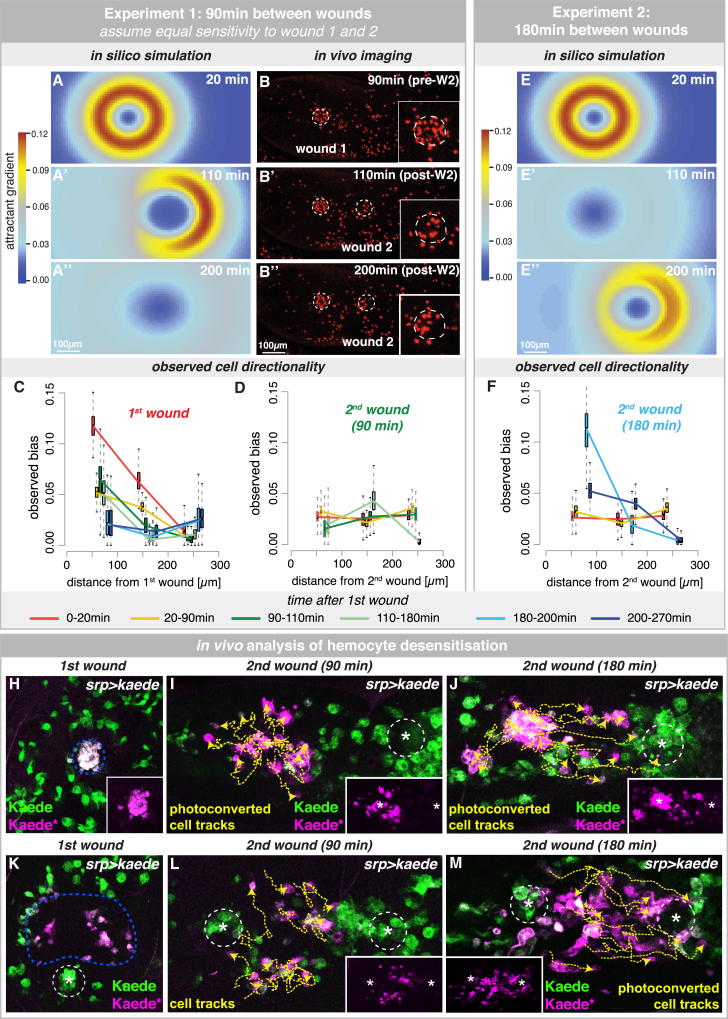
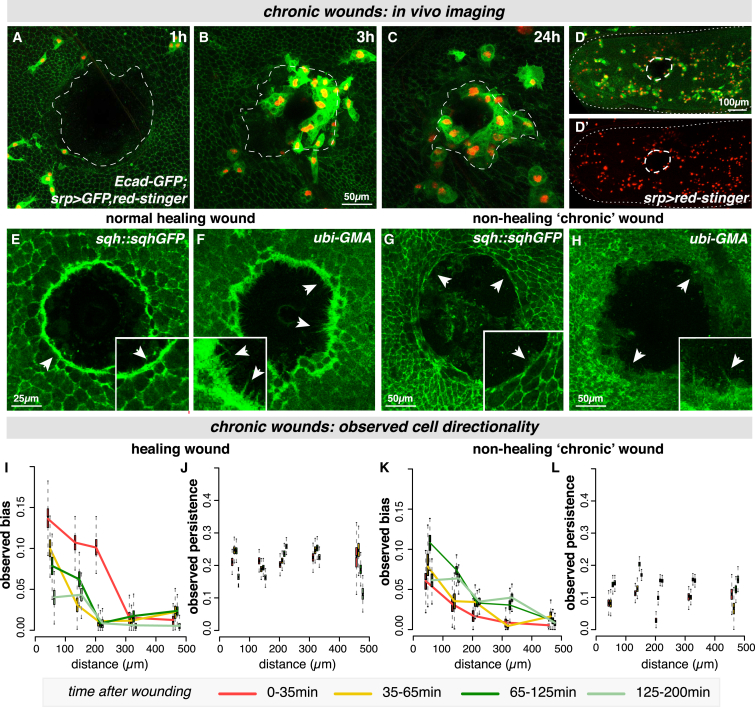
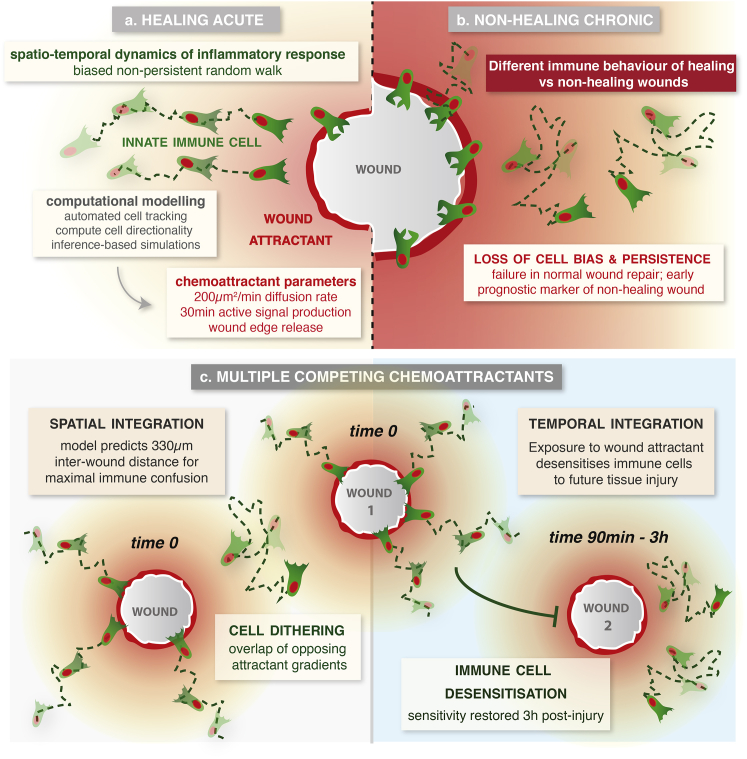
In the acute inflammatory phase following tissue damage, cells of the innate immune system are rapidly recruited to sites of injury by pro-inflammatory mediators released at the wound site. Although advances in live imaging allow us to directly visualize this process in vivo, the precise identity and properties of the primary immune damage attractants remain unclear, as it is currently impossible to directly observe and accurately measure these signals in tissues. Here, we demonstrate that detailed information about the attractant signals can be extracted directly from the in vivo behavior of the responding immune cells. By applying inference-based computational approaches to analyze the in vivo dynamics of the Drosophila inflammatory response, we gain new detailed insight into the spatiotemporal properties of the attractant gradient. In particular, we show that the wound attractant is released by wound margin cells, rather than by the wounded tissue per se, and that it diffuses away from this source at rates far slower than those of previously implicated signals such as H2O2 and ATP, ruling out these fast mediators as the primary chemoattractant. We then predict, and experimentally test, how competing attractant signals might interact in space and time to regulate multi-step cell navigation in the complex environment of a healing wound, revealing a period of receptor desensitization after initial exposure to the damage attractant. Extending our analysis to model much larger wounds, we uncover a dynamic behavioral change in the responding immune cells in vivo that is prognostic of whether a wound will subsequently heal or not. VIDEO ABSTRACT.
Copyright © 2016 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Comment in
-
Wound Signaling: Monkeywrenching Macrophage Migration with Microscopes, Movies, and Math.Curr Biol. 2016 Aug 8;26(15):R715-R717. doi: 10.1016/j.cub.2016.06.023. Curr Biol. 2016. PMID: 27505244
References
-
- Nourshargh S., Hordijk P.L., Sixt M. Breaching multiple barriers: leukocyte motility through venular walls and the interstitium. Nat. Rev. Mol. Cell Biol. 2010;11:366–378. - PubMed
-
- McDonald B., Kubes P. Cellular and molecular choreography of neutrophil recruitment to sites of sterile inflammation. J. Mol. Med. (Berl) 2011;89:1079–1088. - PubMed
-
- McDonald B., Pittman K., Menezes G.B., Hirota S.A., Slaba I., Waterhouse C.C., Beck P.L., Muruve D.A., Kubes P. Intravascular danger signals guide neutrophils to sites of sterile inflammation. Science. 2010;330:362–366. - PubMed
-
- Davalos D., Grutzendler J., Yang G., Kim J.V., Zuo Y., Jung S., Littman D.R., Dustin M.L., Gan W.-B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005;8:752–758. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
