

Origins and Evolution of tetherin, an Orphan Antiviral Gene
- PMID: 27427209
- PMCID: PMC4989275
- DOI: 10.1016/j.chom.2016.06.007
Origins and Evolution of tetherin, an Orphan Antiviral Gene
Abstract
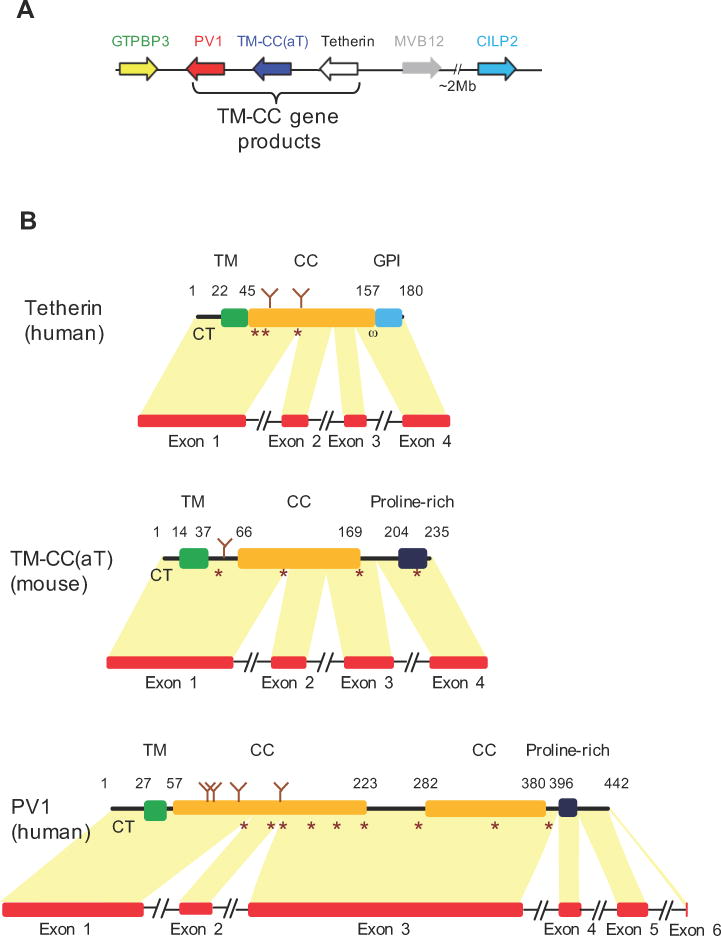
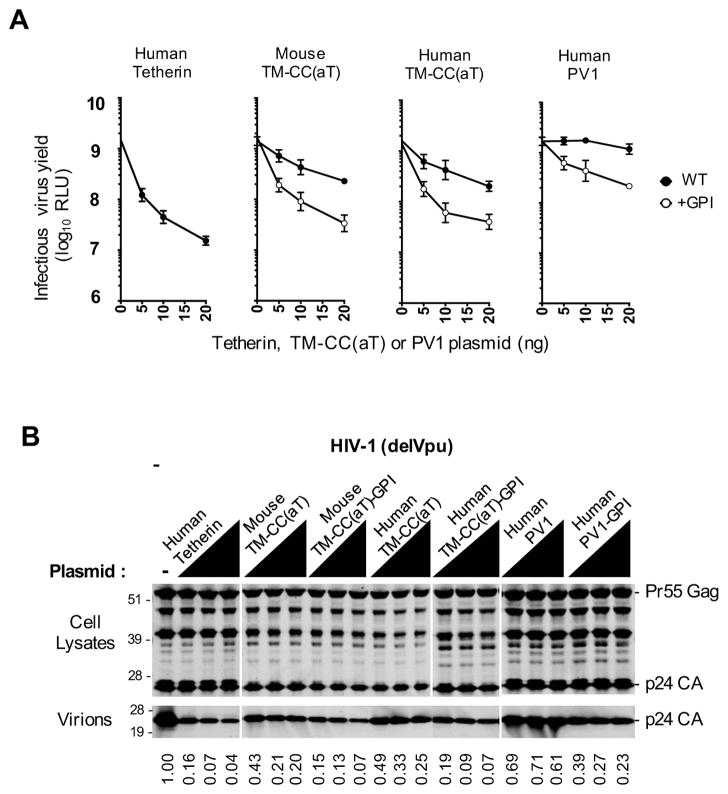
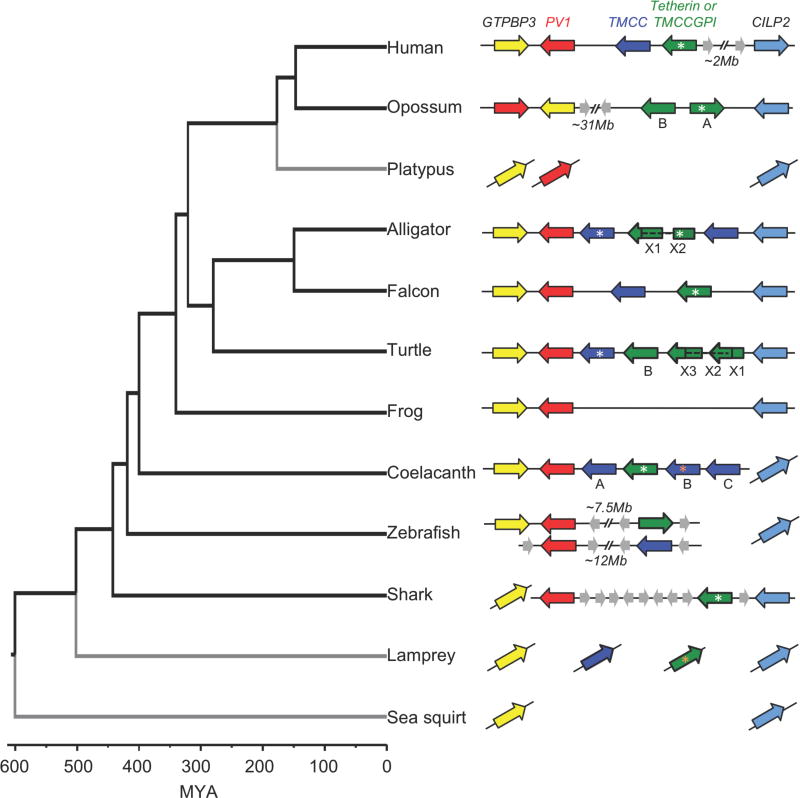
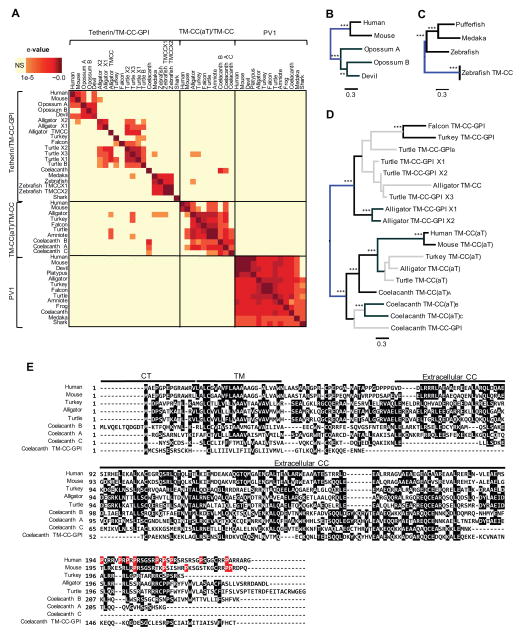
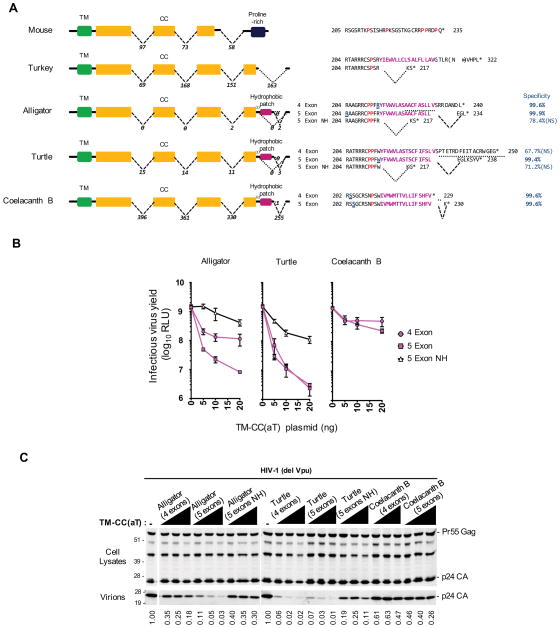
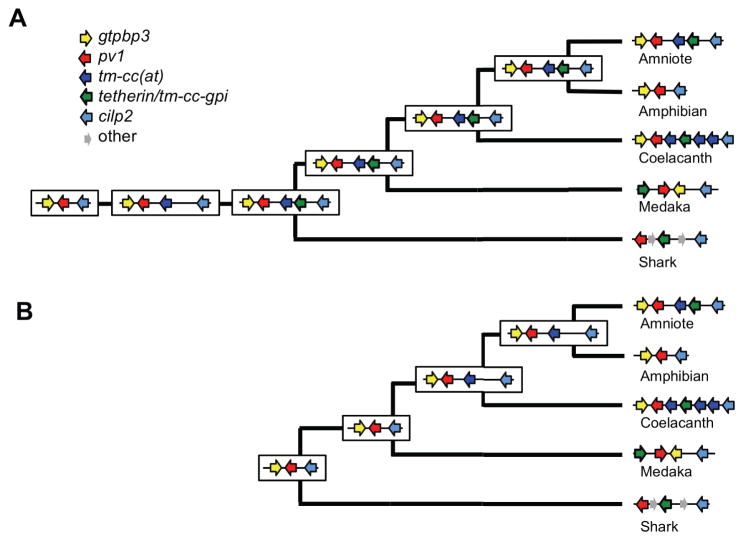
Tetherin encodes an interferon-inducible antiviral protein that traps a broad spectrum of enveloped viruses at infected cell surfaces. Despite the absence of any clearly related gene or activity, we describe possible scenarios by which tetherin arose that exemplify how protein modularity, evolvability, and robustness can create and preserve new functions. We find that tetherin genes in various organisms exhibit no sequence similarity and share only a common architecture and location in modern genomes. Moreover, tetherin is part of a cluster of three potential sister genes encoding proteins of similar architecture, some variants of which exhibit antiviral activity while others can be endowed with antiviral activity by a simple modification. Only in slowly evolving species (e.g., coelacanths) does tetherin exhibit sequence similarity to one potential sister gene. Neofunctionalization, drift, and genetic conflict appear to have driven a near complete loss of sequence similarity among modern tetherin genes and their sister genes.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
