

Infantile amnesia reflects a developmental critical period for hippocampal learning
- PMID: 27428652
- PMCID: PMC5003643
- DOI: 10.1038/nn.4348
Infantile amnesia reflects a developmental critical period for hippocampal learning
Erratum in
-
Erratum: Infantile amnesia reflects a developmental critical period for hippocampal learning.Nat Neurosci. 2017 Jun 27;20(7):1033. doi: 10.1038/nn0717-1033a. Nat Neurosci. 2017. PMID: 28653686 No abstract available.
Abstract
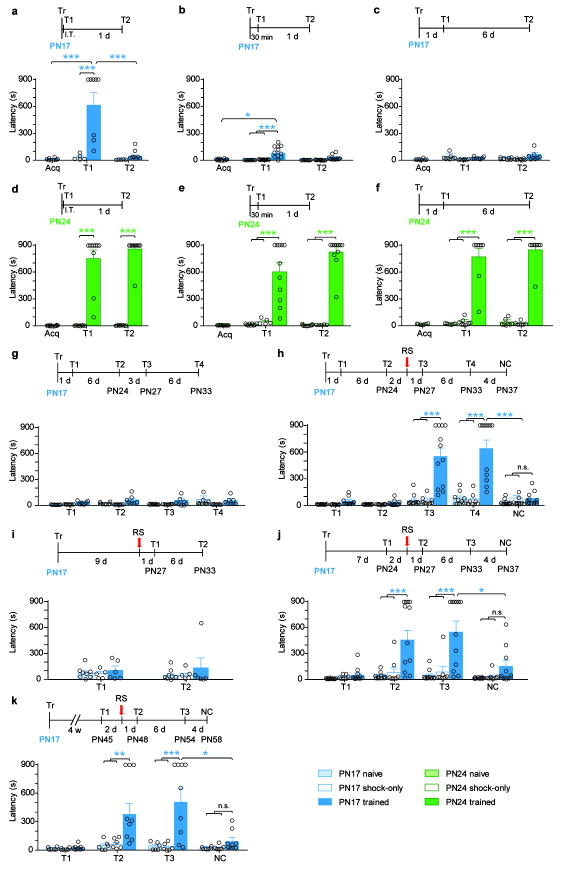
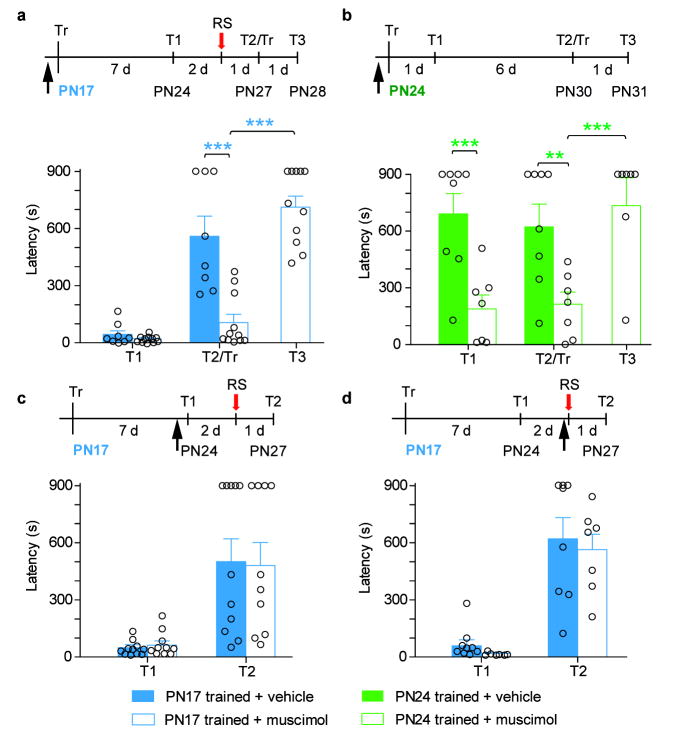
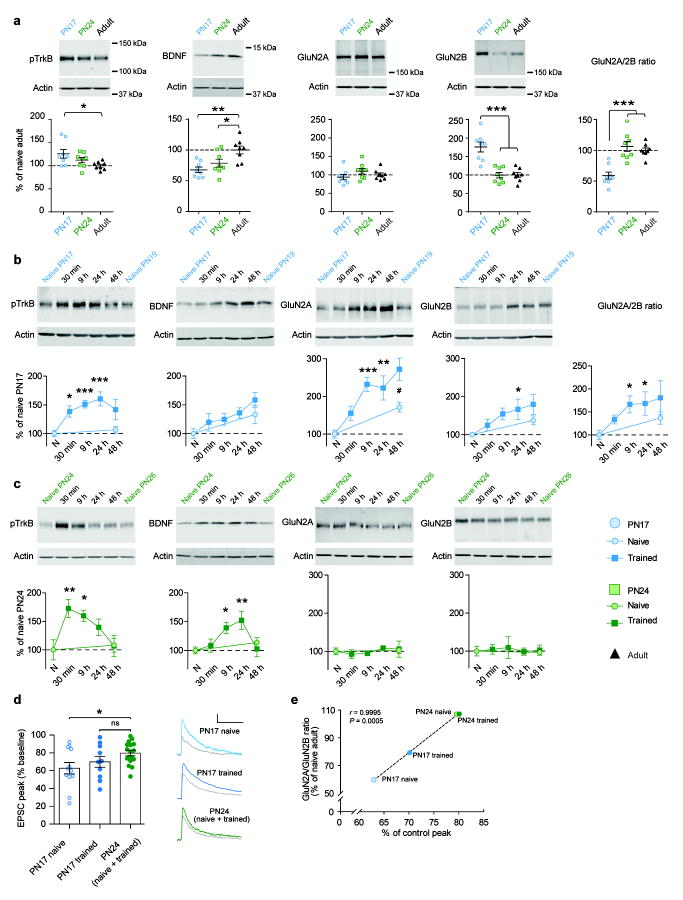
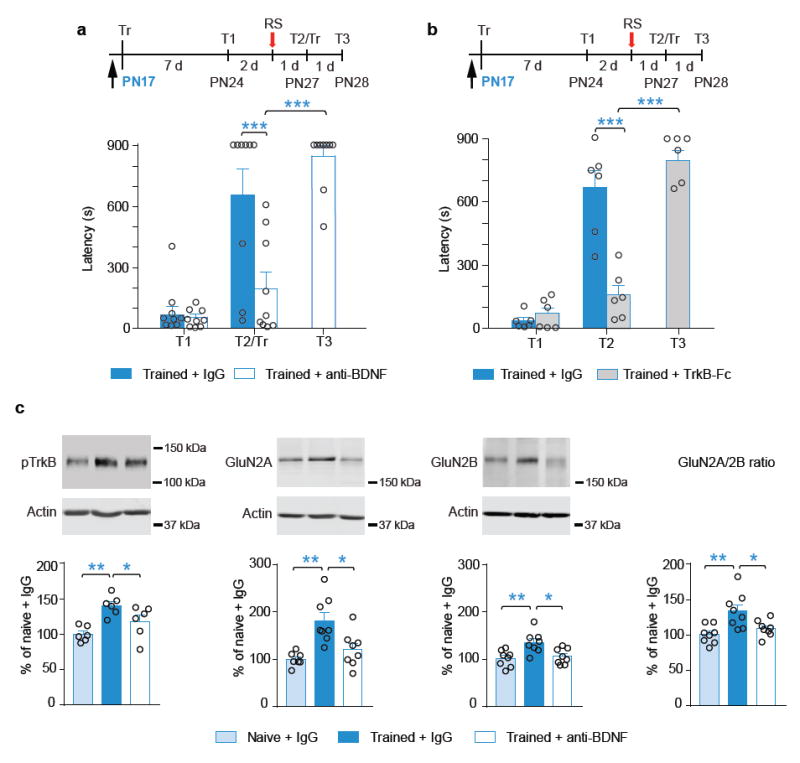
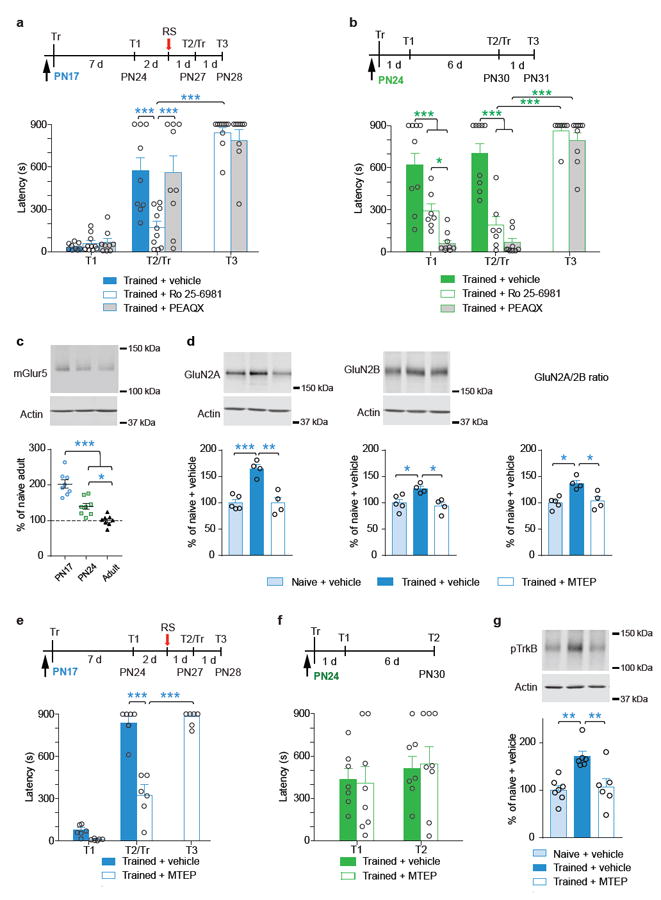
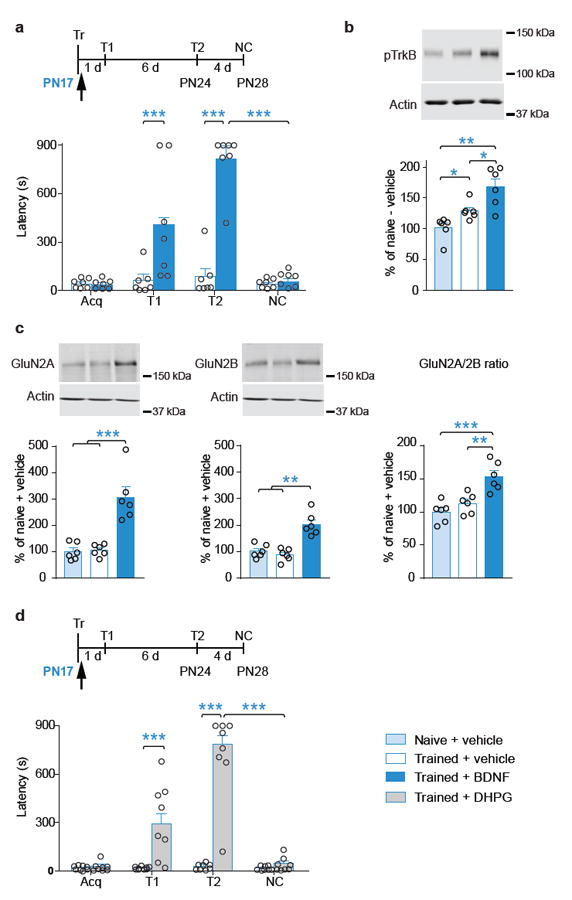
Episodic memories formed during the first postnatal period are rapidly forgotten, a phenomenon known as 'infantile amnesia'. In spite of this memory loss, early experiences influence adult behavior, raising the question of which mechanisms underlie infantile memories and amnesia. Here we show that in rats an experience learned during the infantile amnesia period is stored as a latent memory trace for a long time; indeed, a later reminder reinstates a robust, context-specific and long-lasting memory. The formation and storage of this latent memory requires the hippocampus, follows a sharp temporal boundary and occurs through mechanisms typical of developmental critical periods, including the expression switch of the NMDA receptor subunits from 2B to 2A, which is dependent on brain-derived neurotrophic factor (BDNF) and metabotropic glutamate receptor 5 (mGluR5). Activating BDNF or mGluR5 after training rescues the infantile amnesia. Thus, early episodic memories are not lost but remain stored long term. These data suggest that the hippocampus undergoes a developmental critical period to become functionally competent.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
The hippocampus grows up.Nat Neurosci. 2016 Aug 26;19(9):1190-1. doi: 10.1038/nn.4368. Nat Neurosci. 2016. PMID: 27571198 No abstract available.
References
-
- Campbell BA, Spear NE. Ontogeny of memory. Psychol Rev. 1972;79:215–236. - PubMed
-
- Hayne H. Infant memory development: Implications for childhood amnesia. Dev Rev. 2004;24:33–73.
-
- Rovee-Collier C. The development of infant memory. Curr Dir Psychol Sci. 1999;8:80–85.
-
- Heim C, Nemeroff CB. The role of childhood trauma in the neurobiology of mood and anxiety disorders: preclinical and clinical studies. Biol Psychiatry. 2001;49:1023–1039. - PubMed
-
- Dumas TC, Rudy JW. Development of the hippocampal memory system: Creating networks and modifiable synapses. In: Blumberg MS, Freeman JH, Robinson SR, editors. Oxford Handbook of Developmental Behavioral Neuroscience Oxford library of neuroscience. New York, NY, US: Oxford University Press; 2010.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
