

Perlman syndrome nuclease DIS3L2 controls cytoplasmic non-coding RNAs and provides surveillance pathway for maturing snRNAs
- PMID: 27431325
- PMCID: PMC5137419
- DOI: 10.1093/nar/gkw649
Perlman syndrome nuclease DIS3L2 controls cytoplasmic non-coding RNAs and provides surveillance pathway for maturing snRNAs
Abstract
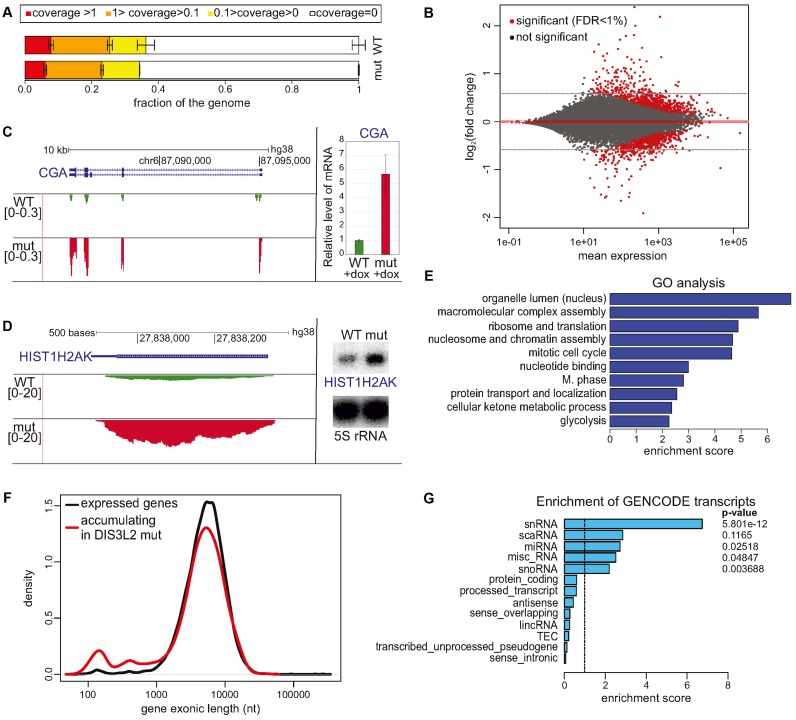
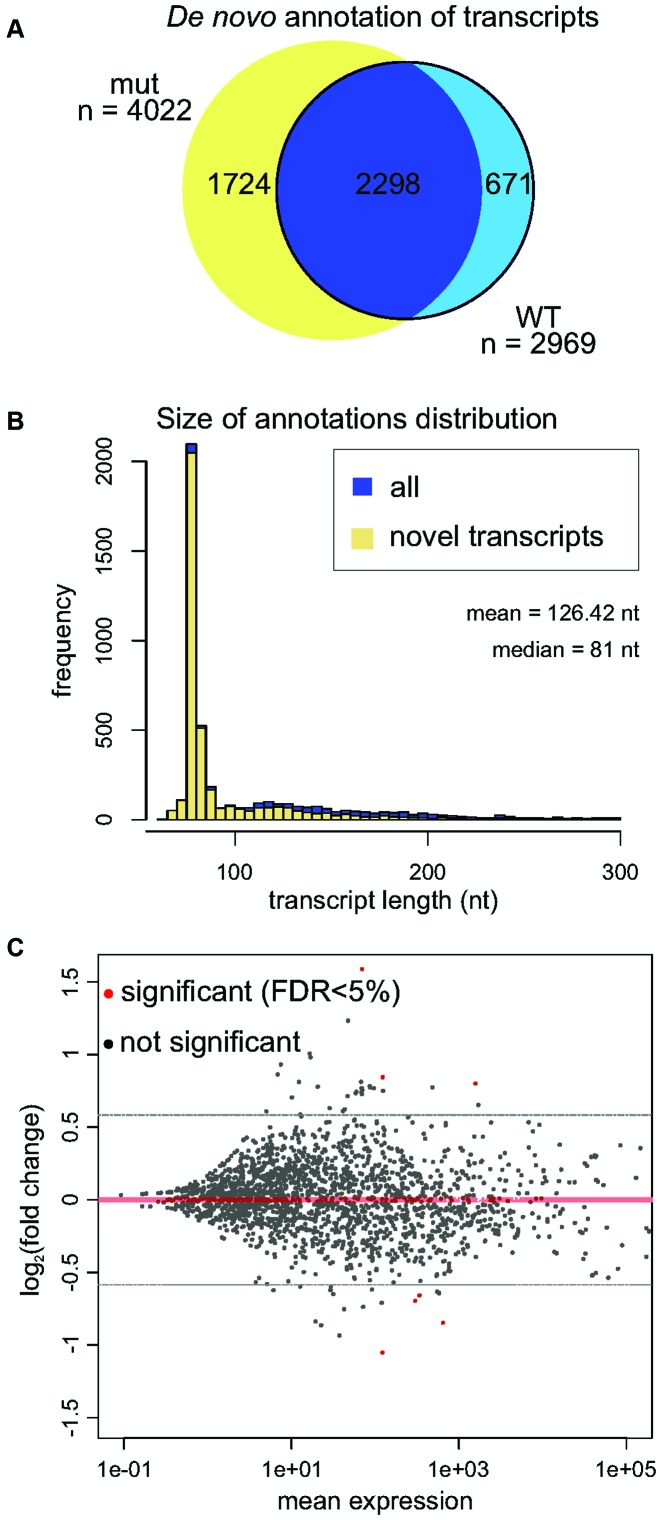
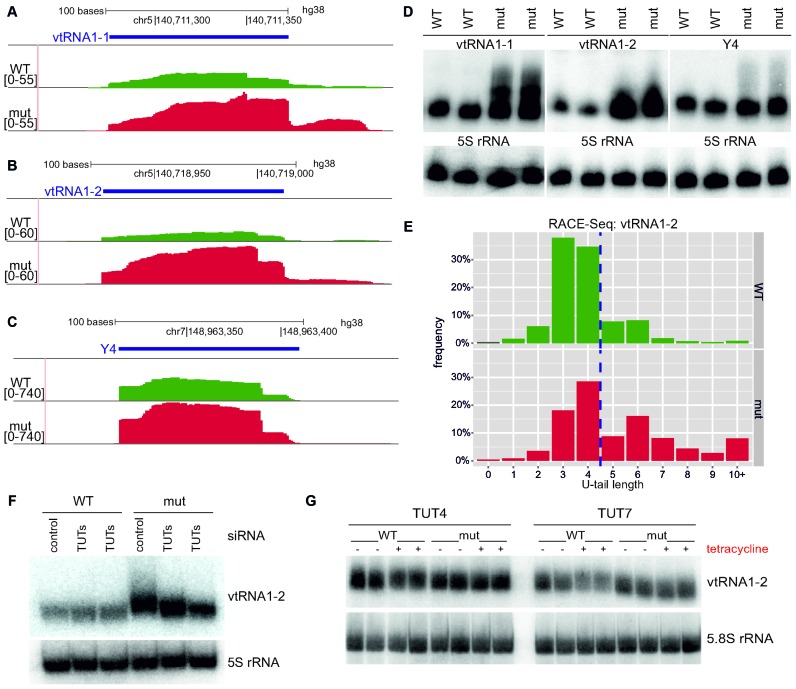
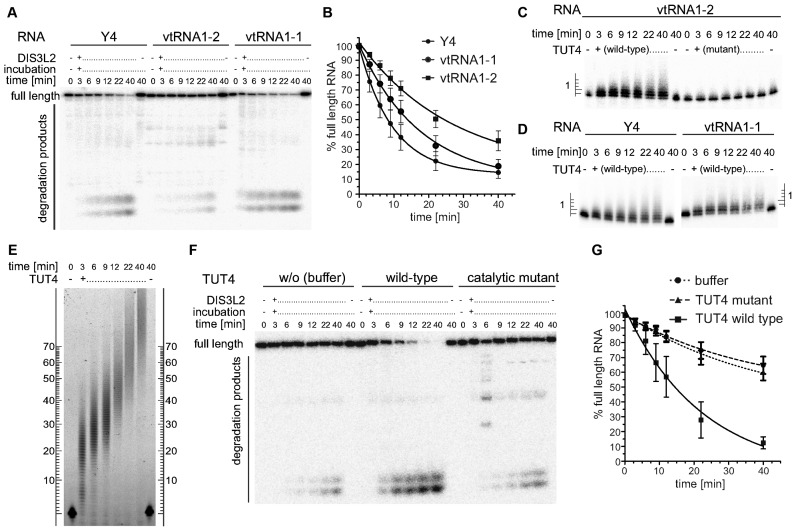
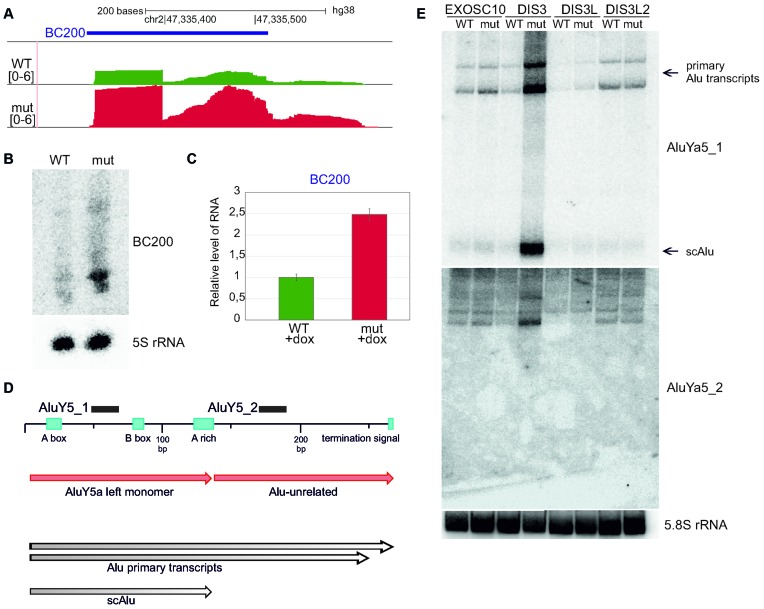
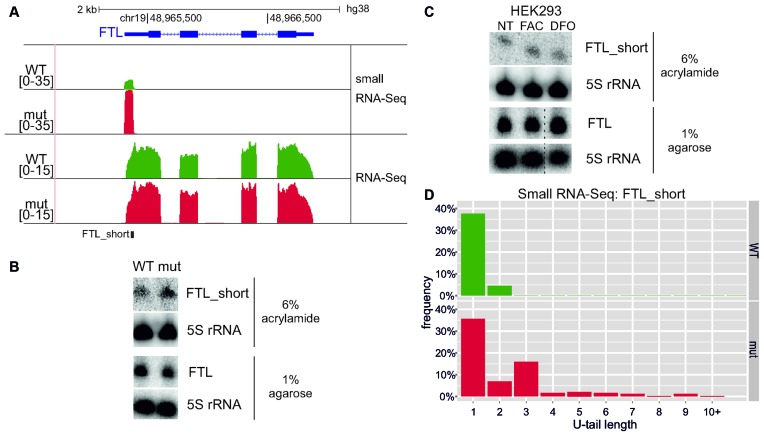
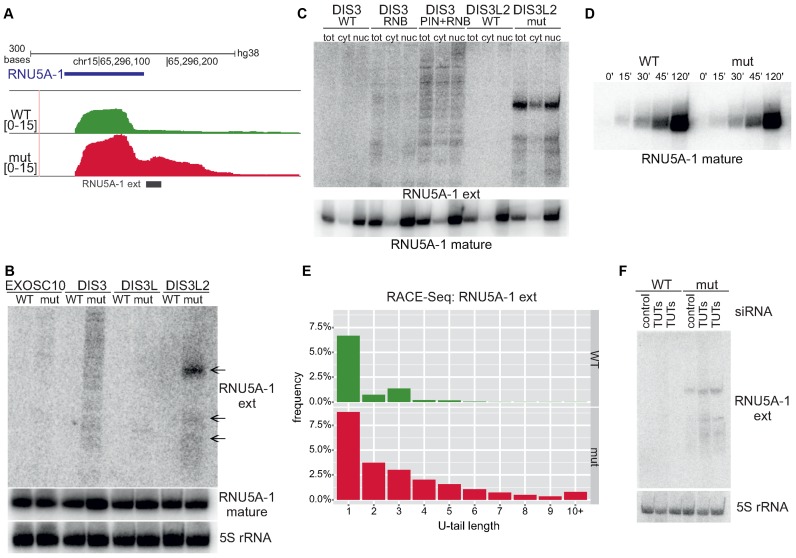
The exosome-independent exoribonuclease DIS3L2 is mutated in Perlman syndrome. Here, we used extensive global transcriptomic and targeted biochemical analyses to identify novel DIS3L2 substrates in human cells. We show that DIS3L2 regulates pol II transcripts, comprising selected canonical and histone-coding mRNAs, and a novel FTL_short RNA from the ferritin mRNA 5' UTR. Importantly, DIS3L2 contributes to surveillance of maturing snRNAs during their cytoplasmic processing. Among pol III transcripts, DIS3L2 particularly targets vault and Y RNAs and an Alu-like element BC200 RNA, but not Alu repeats, which are removed by exosome-associated DIS3. Using 3' RACE-Seq, we demonstrate that all novel DIS3L2 substrates are uridylated in vivo by TUT4/TUT7 poly(U) polymerases. Uridylation-dependent DIS3L2-mediated decay can be recapitulated in vitro, thus reinforcing the tight cooperation between DIS3L2 and TUTases. Together these results indicate that catalytically inactive DIS3L2, characteristic of Perlman syndrome, can lead to deregulation of its target RNAs to disturb transcriptome homeostasis.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
TUT-DIS3L2 is a mammalian surveillance pathway for aberrant structured non-coding RNAs.EMBO J. 2016 Oct 17;35(20):2179-2191. doi: 10.15252/embj.201694857. Epub 2016 Sep 19. EMBO J. 2016. PMID: 27647875 Free PMC article.
-
Dis3l2-Mediated Decay Is a Quality Control Pathway for Noncoding RNAs.Cell Rep. 2016 Aug 16;16(7):1861-73. doi: 10.1016/j.celrep.2016.07.025. Epub 2016 Aug 4. Cell Rep. 2016. PMID: 27498873 Free PMC article.
-
The Perlman syndrome DIS3L2 exoribonuclease safeguards endoplasmic reticulum-targeted mRNA translation and calcium ion homeostasis.Nat Commun. 2020 May 26;11(1):2619. doi: 10.1038/s41467-020-16418-y. Nat Commun. 2020. PMID: 32457326 Free PMC article.
-
Regulation of RNA decay and cellular function by 3'-5' exoribonuclease DIS3L2.RNA Biol. 2019 Feb;16(2):160-165. doi: 10.1080/15476286.2018.1564466. Epub 2019 Jan 13. RNA Biol. 2019. PMID: 30638126 Free PMC article. Review.
-
Unraveling 3'-end RNA uridylation at nucleotide resolution.Methods. 2019 Feb 15;155:10-19. doi: 10.1016/j.ymeth.2018.10.024. Epub 2018 Nov 3. Methods. 2019. PMID: 30395968 Free PMC article. Review.
Cited by
-
RNA uridylation and decay in plants.Philos Trans R Soc Lond B Biol Sci. 2018 Nov 5;373(1762):20180163. doi: 10.1098/rstb.2018.0163. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30397100 Free PMC article. Review.
-
DIS3L2 and LSm proteins are involved in the surveillance of Sm ring-deficient snRNAs.Nucleic Acids Res. 2020 Jun 19;48(11):6184-6197. doi: 10.1093/nar/gkaa301. Nucleic Acids Res. 2020. PMID: 32374871 Free PMC article.
-
RNA Metabolism Guided by RNA Modifications: The Role of SMUG1 in rRNA Quality Control.Biomolecules. 2021 Jan 8;11(1):76. doi: 10.3390/biom11010076. Biomolecules. 2021. PMID: 33430019 Free PMC article. Review.
-
Cytoplasmic mRNA decay and quality control machineries in eukaryotes.Nat Rev Genet. 2025 Jul;26(7):463-478. doi: 10.1038/s41576-024-00810-1. Epub 2025 Jan 27. Nat Rev Genet. 2025. PMID: 39870755 Review.
-
LINE-1 mRNA 3' end dynamics shape its biology and retrotransposition potential.Nucleic Acids Res. 2024 Apr 12;52(6):3327-3345. doi: 10.1093/nar/gkad1251. Nucleic Acids Res. 2024. PMID: 38197223 Free PMC article.
References
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
