

'Biogeneric' developmental processes: drivers of major transitions in animal evolution
- PMID: 27431521
- PMCID: PMC4958937
- DOI: 10.1098/rstb.2015.0443
'Biogeneric' developmental processes: drivers of major transitions in animal evolution
Abstract
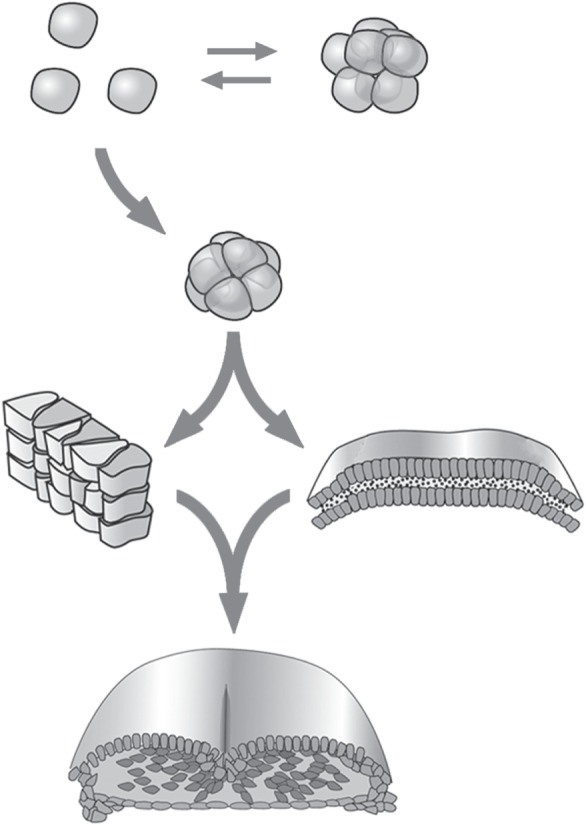
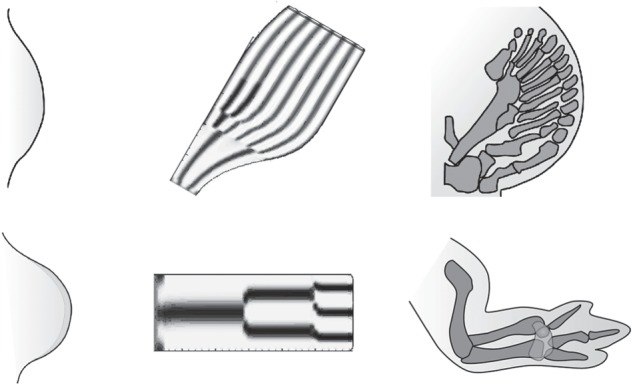
Using three examples drawn from animal systems, I advance the hypothesis that major transitions in multicellular evolution often involved the constitution of new cell-based materials with unprecedented morphogenetic capabilities. I term the materials and formative processes that arise when highly evolved cells are incorporated into mesoscale matter 'biogeneric', to reflect their commonality with, and distinctiveness from, the organizational properties of non-living materials. The first transition arose by the innovation of classical cell-adhesive cadherins with transmembrane linkage to the cytoskeleton and the appearance of the morphogen Wnt, transforming some ancestral unicellular holozoans into 'liquid tissues', and thereby originating the metazoans. The second transition involved the new capabilities, within a basal metazoan population, of producing a mechanically stable basal lamina, and of planar cell polarization. This gave rise to the eumetazoans, initially diploblastic (two-layered) forms, and then with the addition of extracellular matrices promoting epithelial-mesenchymal transformation, three-layered triploblasts. The last example is the fin-to-limb transition. Here, the components of a molecular network that promoted the development of species-idiosyncratic endoskeletal elements in gnathostome ancestors are proposed to have evolved to a dynamical regime in which they constituted a Turing-type reaction-diffusion system capable of organizing the stereotypical arrays of elements of lobe-finned fish and tetrapods. The contrasting implications of the biogeneric materials-based and neo-Darwinian perspectives for understanding major evolutionary transitions are discussed.This article is part of the themed issue 'The major synthetic evolutionary transitions'.
Keywords: diploblasty; liquid tissues; morphological evolution; reaction–diffusion mechanism; tetrapod limb; triploblasty.
© 2016 The Author(s).
Figures
Similar articles
-
The vertebrate limb: An evolving complex of self-organizing systems.Prog Biophys Mol Biol. 2018 Sep;137:12-24. doi: 10.1016/j.pbiomolbio.2018.01.002. Epub 2018 Jan 8. Prog Biophys Mol Biol. 2018. PMID: 29325895 Review.
-
Ancestral developmental potentials in early bony fish contributed to vertebrate water-to-land transition.Zool Res. 2021 Mar 18;42(2):135-137. doi: 10.24272/j.issn.2095-8137.2021.066. Zool Res. 2021. PMID: 33709637 Free PMC article.
-
Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins-to-limbs transition.Biol Rev Camb Philos Soc. 2018 May;93(2):1077-1107. doi: 10.1111/brv.12386. Epub 2017 Nov 10. Biol Rev Camb Philos Soc. 2018. PMID: 29125205 Review.
-
Characteristic tetrapod musculoskeletal limb phenotype emerged more than 400 MYA in basal lobe-finned fishes.Sci Rep. 2016 Nov 25;6:37592. doi: 10.1038/srep37592. Sci Rep. 2016. PMID: 27886207 Free PMC article.
-
A Devonian tetrapod-like fish and the evolution of the tetrapod body plan.Nature. 2006 Apr 6;440(7085):757-63. doi: 10.1038/nature04639. Nature. 2006. PMID: 16598249
Cited by
-
Inherency of Form and Function in Animal Development and Evolution.Front Physiol. 2019 Jun 19;10:702. doi: 10.3389/fphys.2019.00702. eCollection 2019. Front Physiol. 2019. PMID: 31275153 Free PMC article. Review.
-
Why call it developmental bias when it is just development?Biol Direct. 2021 Jan 9;16(1):3. doi: 10.1186/s13062-020-00289-w. Biol Direct. 2021. PMID: 33422150 Free PMC article.
-
Dynamical Patterning Modules, Biogeneric Materials, and the Evolution of Multicellular Plants.Front Plant Sci. 2018 Jul 16;9:871. doi: 10.3389/fpls.2018.00871. eCollection 2018. Front Plant Sci. 2018. PMID: 30061903 Free PMC article.
-
Interplay of mesoscale physics and agent-like behaviors in the parallel evolution of aggregative multicellularity.Evodevo. 2020 Oct 12;11:21. doi: 10.1186/s13227-020-00165-8. eCollection 2020. Evodevo. 2020. PMID: 33062243 Free PMC article. Review.
-
Living tissues are more than cell clusters: The extracellular matrix as a driving force in morphogenesis.Prog Biophys Mol Biol. 2018 Sep;137:46-51. doi: 10.1016/j.pbiomolbio.2018.01.009. Epub 2018 Jan 31. Prog Biophys Mol Biol. 2018. PMID: 29398066 Free PMC article. Review.
References
-
- Nielsen C. 2012. Animal evolution: interrelationships of the living phyla, 3rd edn Oxford, UK: Oxford University Press.
-
- Forgacs G, Newman SA. 2005. Biological physics of the developing embryo. Cambridge, UK: Cambridge University Press.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
