

Stabilizing multicellularity through ratcheting
- PMID: 27431522
- PMCID: PMC4958938
- DOI: 10.1098/rstb.2015.0444
Stabilizing multicellularity through ratcheting
Abstract
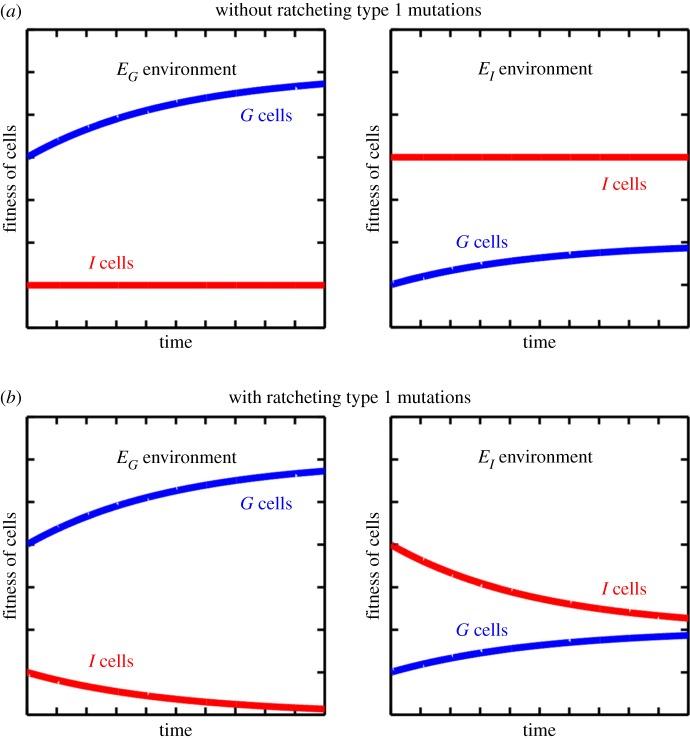
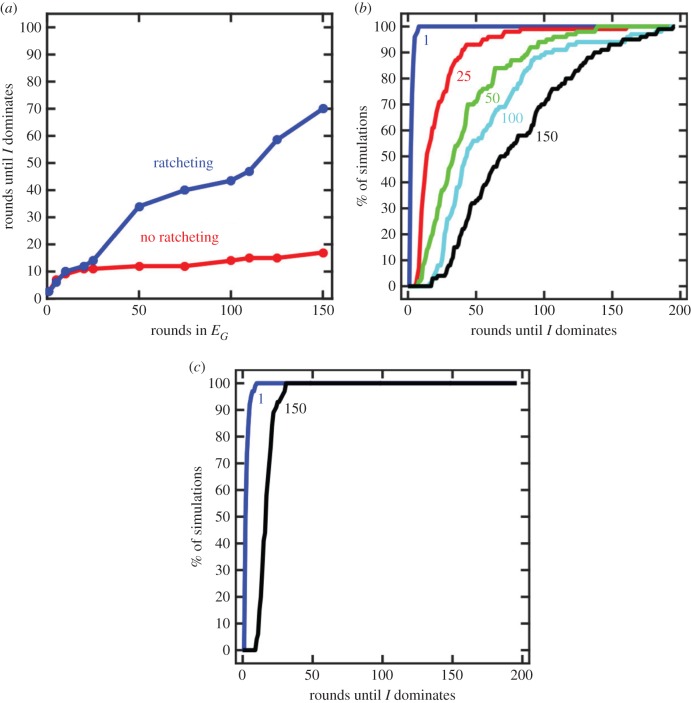
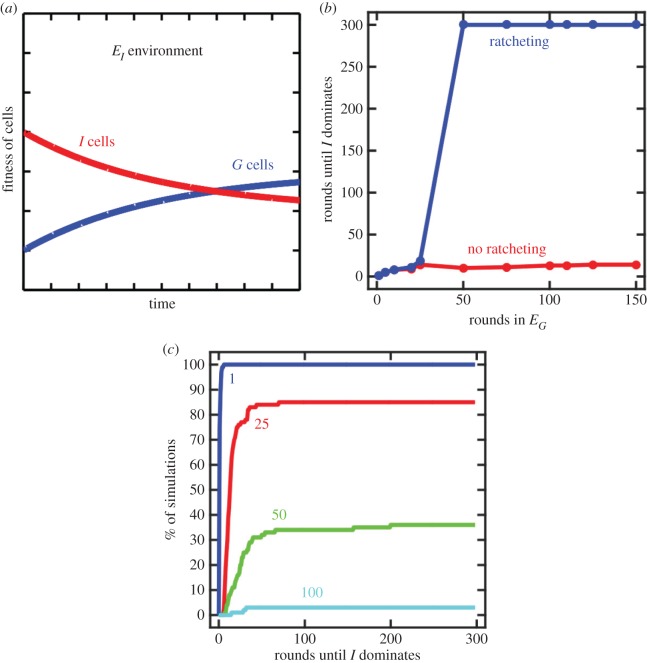
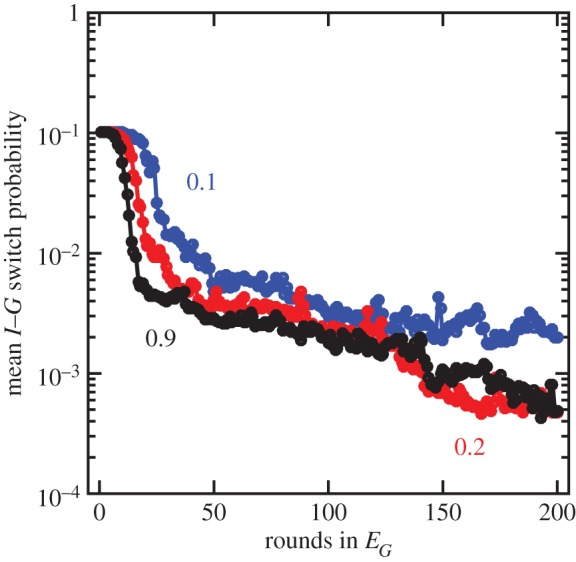
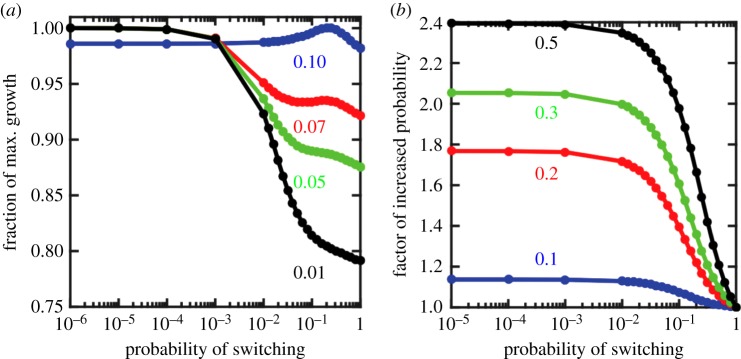
The evolutionary transition to multicellularity probably began with the formation of simple undifferentiated cellular groups. Such groups evolve readily in diverse lineages of extant unicellular taxa, suggesting that there are few genetic barriers to this first key step. This may act as a double-edged sword: labile transitions between unicellular and multicellular states may facilitate the evolution of simple multicellularity, but reversion to a unicellular state may inhibit the evolution of increased complexity. In this paper, we examine how multicellular adaptations can act as evolutionary 'ratchets', limiting the potential for reversion to unicellularity. We consider a nascent multicellular lineage growing in an environment that varies between favouring multicellularity and favouring unicellularity. The first type of ratcheting mutations increase cell-level fitness in a multicellular context but are costly in a single-celled context, reducing the fitness of revertants. The second type of ratcheting mutations directly decrease the probability that a mutation will result in reversion (either as a pleiotropic consequence or via direct modification of switch rates). We show that both types of ratcheting mutations act to stabilize the multicellular state. We also identify synergistic effects between the two types of ratcheting mutations in which the presence of one creates the selective conditions favouring the other. Ratcheting mutations may play a key role in diverse evolutionary transitions in individuality, sustaining selection on the new higher-level organism by constraining evolutionary reversion.This article is part of the themed issue 'The major synthetic evolutionary transitions'.
Keywords: evolution; major transition; multicellularity; ratcheting; stability.
© 2016 The Authors.
Figures
References
-
- Buss LW. 1987. The evolution of individuality. Princeton, NJ: Princeton University Press.
-
- Smith JM, Szathmáry E. 1995. The major transitions in evolution. Oxford, UK: Oxford University Press.
-
- Michod RE, Roze D. 1999. Cooperation and conflict in the evolution of individuality. III. Transitions in the unit of fitness. In Mathematical and computational biology: computational morphogenesis, hierarchical complexity, and digital evolution (ed. Nehaniv CL.), pp. 47–92. Providence, RI: American Mathematical Society.
-
- Grosberg RK, Strathmann RR. 2007. The evolution of multicellularity: a minor major transition? Annu. Rev. Ecol. Evol. Syst. 38, 621–654. (10.1146/annurev.ecolsys.36.102403.114735) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
