

Synthetic consciousness: the distributed adaptive control perspective
- PMID: 27431526
- PMCID: PMC4958942
- DOI: 10.1098/rstb.2015.0448
Synthetic consciousness: the distributed adaptive control perspective
Abstract
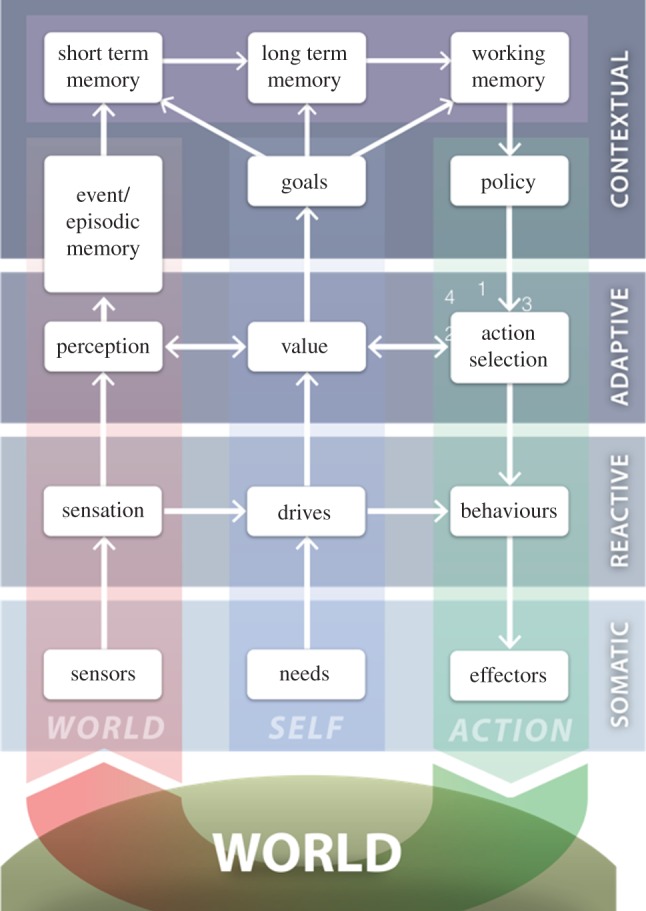
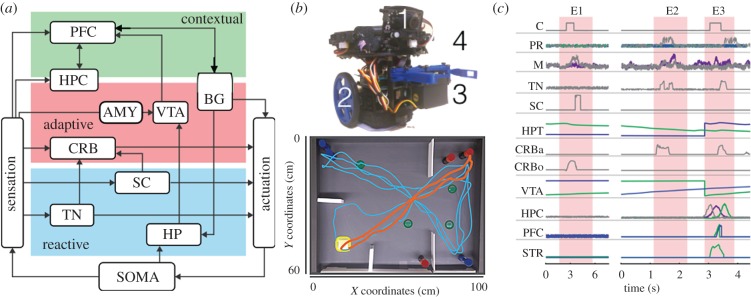
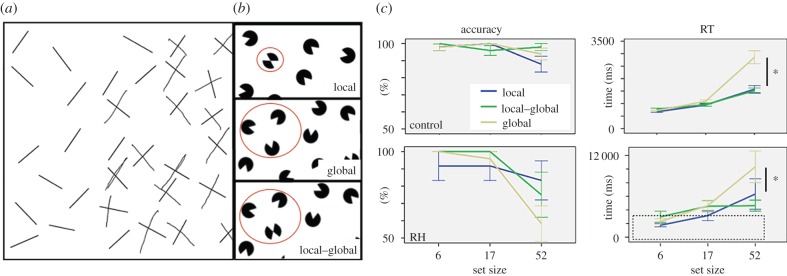
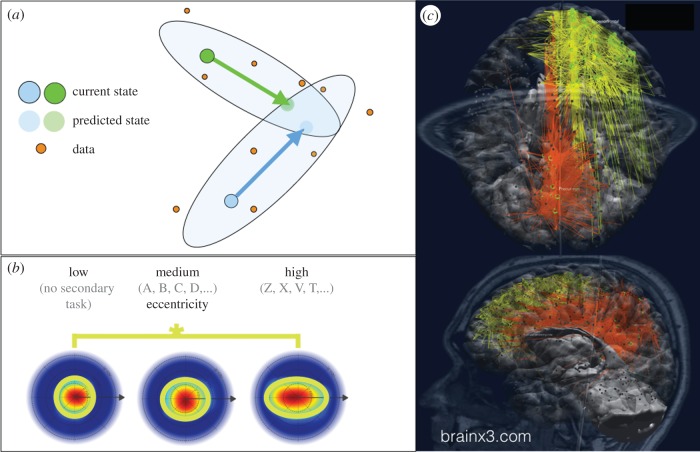
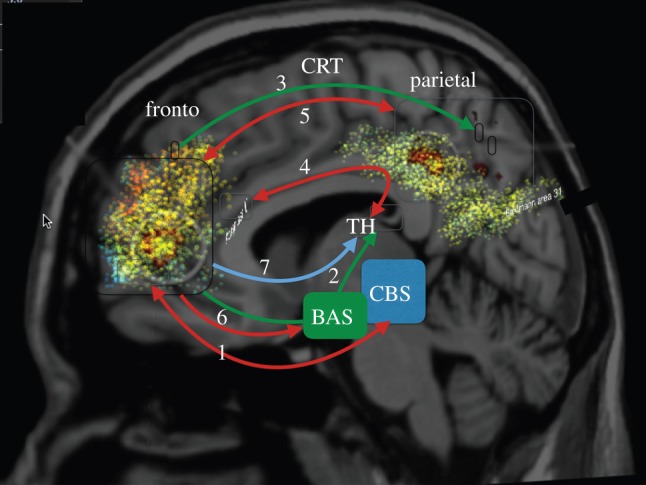
Understanding the nature of consciousness is one of the grand outstanding scientific challenges. The fundamental methodological problem is how phenomenal first person experience can be accounted for in a third person verifiable form, while the conceptual challenge is to both define its function and physical realization. The distributed adaptive control theory of consciousness (DACtoc) proposes answers to these three challenges. The methodological challenge is answered relative to the hard problem and DACtoc proposes that it can be addressed using a convergent synthetic methodology using the analysis of synthetic biologically grounded agents, or quale parsing. DACtoc hypothesizes that consciousness in both its primary and secondary forms serves the ability to deal with the hidden states of the world and emerged during the Cambrian period, affording stable multi-agent environments to emerge. The process of consciousness is an autonomous virtualization memory, which serializes and unifies the parallel and subconscious simulations of the hidden states of the world that are largely due to other agents and the self with the objective to extract norms. These norms are in turn projected as value onto the parallel simulation and control systems that are driving action. This functional hypothesis is mapped onto the brainstem, midbrain and the thalamo-cortical and cortico-cortical systems and analysed with respect to our understanding of deficits of consciousness. Subsequently, some of the implications and predictions of DACtoc are outlined, in particular, the prediction that normative bootstrapping of conscious agents is predicated on an intentionality prior. In the view advanced here, human consciousness constitutes the ultimate evolutionary transition by allowing agents to become autonomous with respect to their evolutionary priors leading to a post-biological Anthropocene.This article is part of the themed issue 'The major synthetic evolutionary transitions'.
Keywords: consciousness; distributed adaptive control; memory; neuronal substrate; normativity; social brain.
© 2016 The Author(s).
Figures
Similar articles
-
Consciousness, biology and quantum hypotheses.Phys Life Rev. 2012 Sep;9(3):285-94. doi: 10.1016/j.plrev.2012.07.001. Epub 2012 Jul 10. Phys Life Rev. 2012. PMID: 22925839 Review.
-
An Integrated World Modeling Theory (IWMT) of Consciousness: Combining Integrated Information and Global Neuronal Workspace Theories With the Free Energy Principle and Active Inference Framework; Toward Solving the Hard Problem and Characterizing Agentic Causation.Front Artif Intell. 2020 Jun 9;3:30. doi: 10.3389/frai.2020.00030. eCollection 2020. Front Artif Intell. 2020. PMID: 33733149 Free PMC article.
-
Towards solving the hard problem of consciousness: The varieties of brain resonances and the conscious experiences that they support.Neural Netw. 2017 Mar;87:38-95. doi: 10.1016/j.neunet.2016.11.003. Epub 2016 Dec 6. Neural Netw. 2017. PMID: 28088645 Review.
-
From the origins to the stream of consciousness and its neural correlates.Front Integr Neurosci. 2022 Nov 4;16:928978. doi: 10.3389/fnint.2022.928978. eCollection 2022. Front Integr Neurosci. 2022. PMID: 36407293 Free PMC article.
-
Genomic instantiation of consciousness in neurons through a biophoton field theory.J Integr Neurosci. 2014 Jun;13(2):253-92. doi: 10.1142/S0219635214400081. Epub 2014 Jul 4. J Integr Neurosci. 2014. PMID: 25012712
Cited by
-
Epistemic Autonomy: Self-supervised Learning in the Mammalian Hippocampus.Trends Cogn Sci. 2021 Jul;25(7):582-595. doi: 10.1016/j.tics.2021.03.016. Epub 2021 Apr 24. Trends Cogn Sci. 2021. PMID: 33906817 Free PMC article. Review.
-
Distinguishing Self, Other, and Autonomy From Visual Feedback: A Combined Correlation and Acceleration Transfer Analysis.Front Hum Neurosci. 2021 Sep 1;15:560657. doi: 10.3389/fnhum.2021.560657. eCollection 2021. Front Hum Neurosci. 2021. PMID: 34539361 Free PMC article.
-
The Morphospace of Consciousness: Three Kinds of Complexity for Minds and Machines.NeuroSci. 2023 Mar 27;4(2):79-102. doi: 10.3390/neurosci4020009. eCollection 2023 Jun. NeuroSci. 2023. PMID: 39483317 Free PMC article.
-
'Seeing the Dark': Grounding Phenomenal Transparency and Opacity in Precision Estimation for Active Inference.Front Psychol. 2018 May 4;9:643. doi: 10.3389/fpsyg.2018.00643. eCollection 2018. Front Psychol. 2018. PMID: 29780343 Free PMC article.
-
Modeling the formation of social conventions from embodied real-time interactions.PLoS One. 2020 Jun 22;15(6):e0234434. doi: 10.1371/journal.pone.0234434. eCollection 2020. PLoS One. 2020. PMID: 32569266 Free PMC article.
References
-
- Crick F, Koch C. 1990. Towards a neurobiological theory of consciousness. Semin. Neurosci. 2, 263–275. (10.1016/B978-0-12-185254-2.50021-8) - DOI
-
- Edelman GM. 1989. The remembered present: a biological theory of consciousness. New York, NY: Basic Books.
-
- Finkelstein G. 2014. Emil du Bois-Reymond‘s reflections on consciousness. In Brain, mind and consciousness in the history of neuroscience (eds Smith CUM, Whitaker H), pp. 163–184. Dordrecht, The Netherlands: Springer.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
