

Adeno Associated Virus 9-Based Gene Therapy Delivers a Functional Monocarboxylate Transporter 8, Improving Thyroid Hormone Availability to the Brain of Mct8-Deficient Mice
- PMID: 27432638
- PMCID: PMC5036314
- DOI: 10.1089/thy.2016.0060
Adeno Associated Virus 9-Based Gene Therapy Delivers a Functional Monocarboxylate Transporter 8, Improving Thyroid Hormone Availability to the Brain of Mct8-Deficient Mice
Abstract
Background: MCT8 gene mutations produce thyroid hormone (TH) deficiency in the brain, causing severe neuropsychomotor abnormalities not correctable by treatment with TH. This proof-of-concept study examined whether transfer of human MCT8 (hMCT8) cDNA using adeno-associated virus 9 (AAV9) could correct the brain defects of Mct8 knockout mice (Mct8KO).
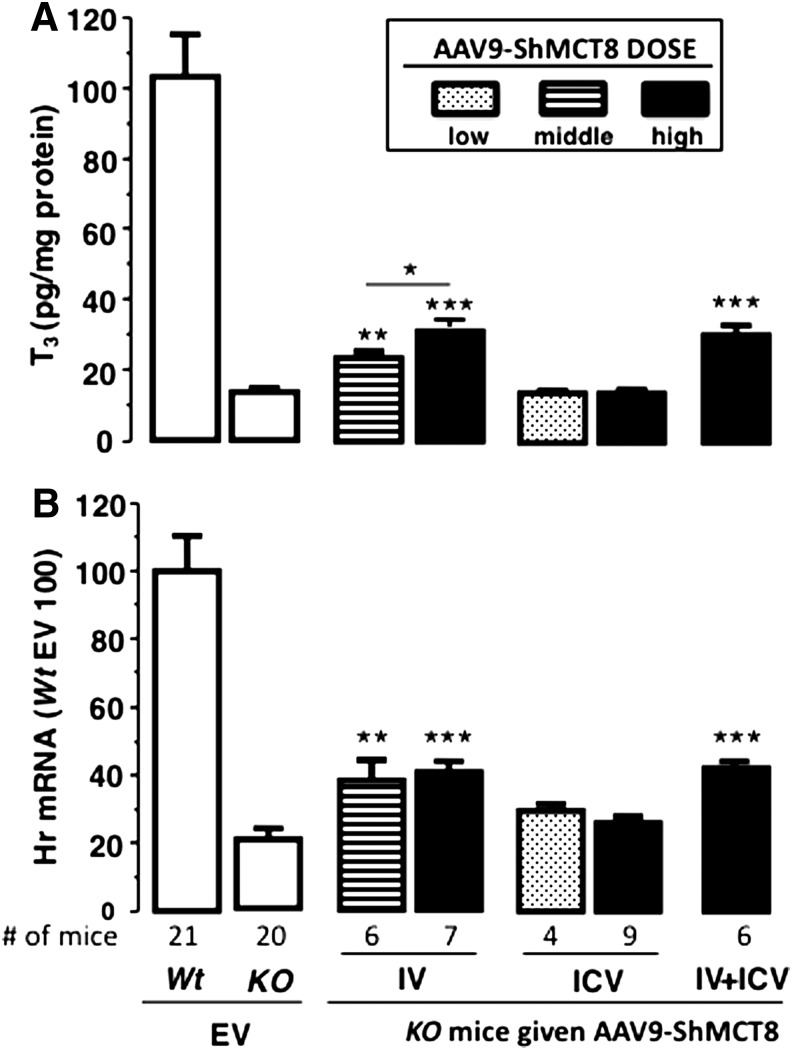
Methods: AAV9 vectors delivering long and/or short hMCT8 protein isoforms or an empty vector were injected intravenously (IV) and/or intracerebroventricularly (ICV) into postnatal day 1 Mct8KO and wild type (Wt) mice. Triiodothyronine (T3) was given daily for four days before postnatal day 28, at which time brains were collected after perfusion to assess increase in T3 content and effect on the T3-responsive transcription factor, Hairless.
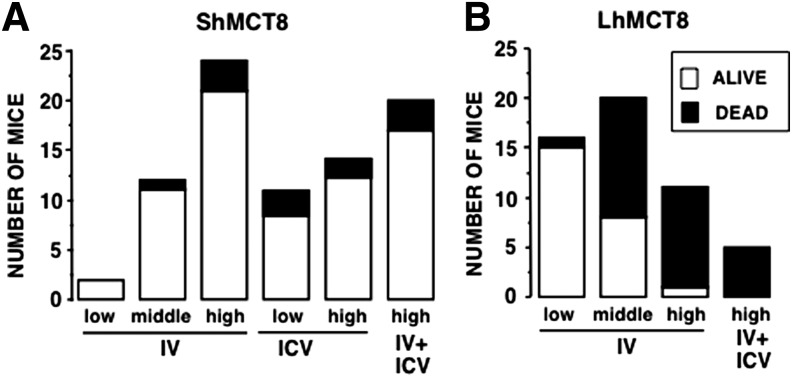
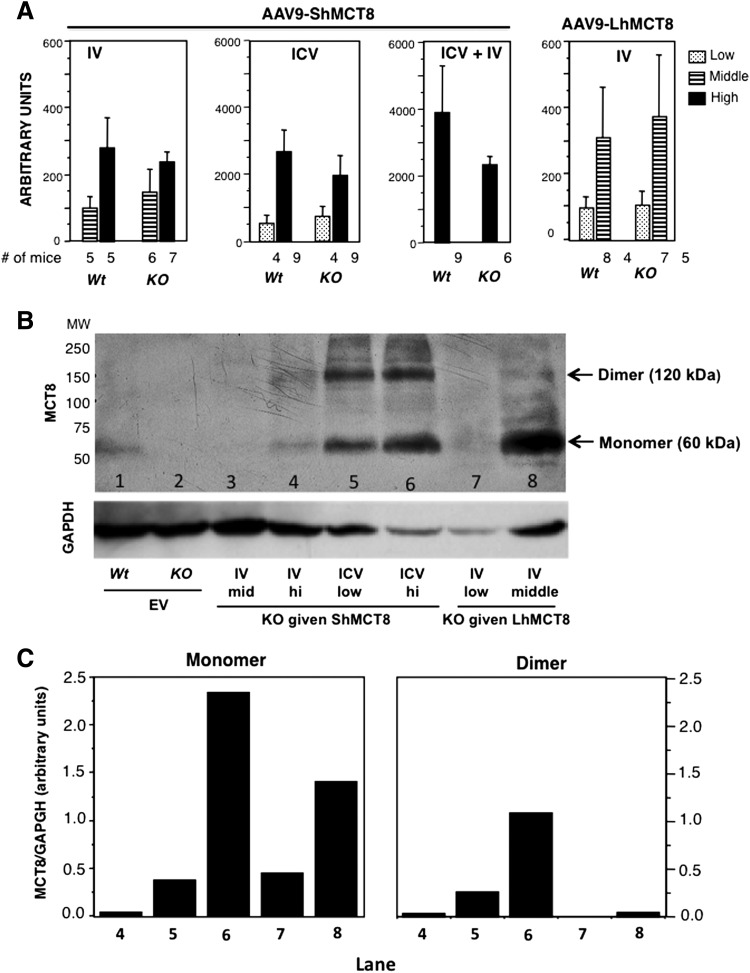
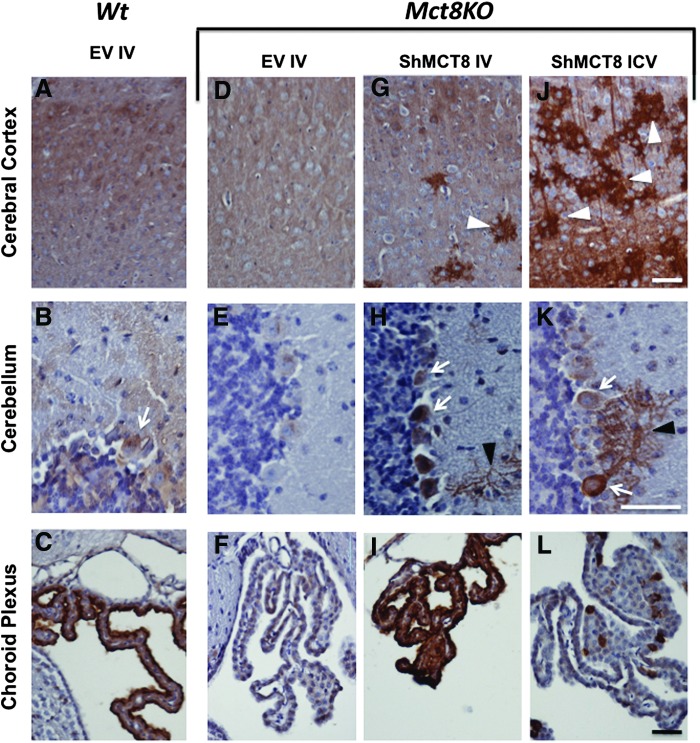
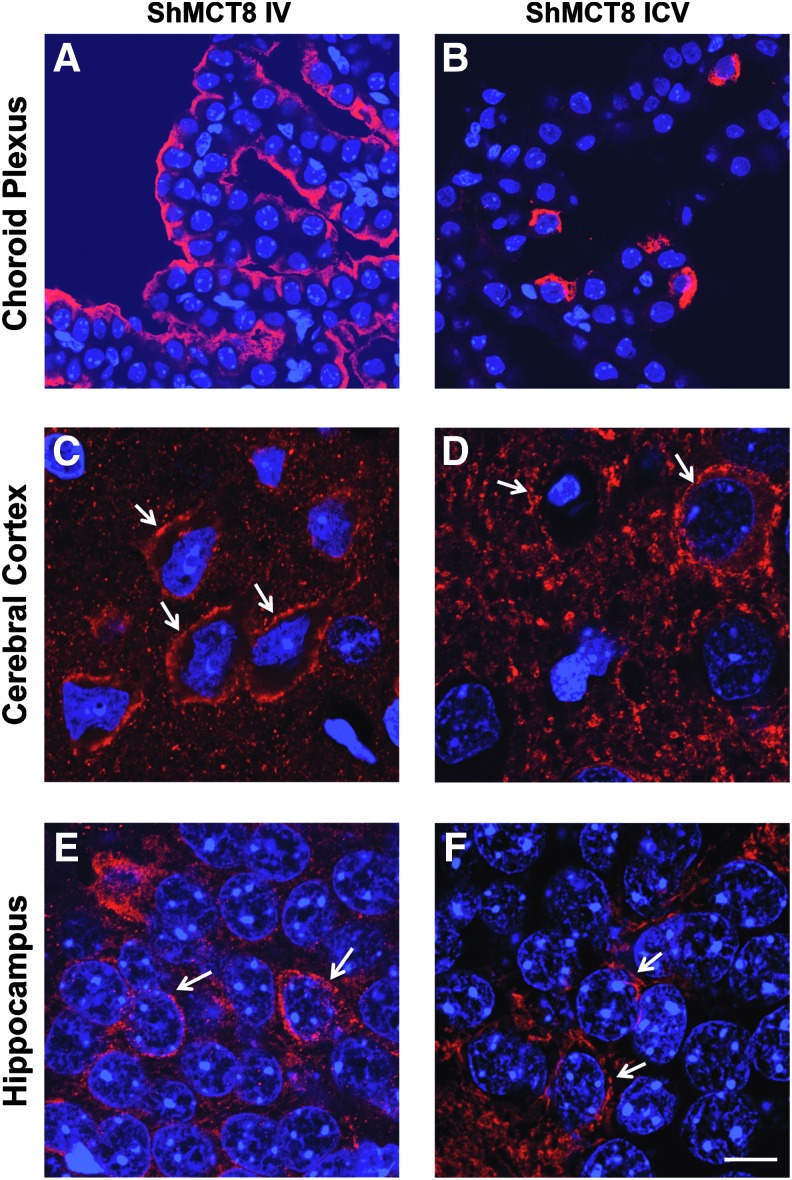
Results: Increased pup mortality was observed after IV injection of the AAV9-long hMCT8 isoform, but not after injection of AAV9-short hMCT8 isoform. Compared to IV, ICV delivery produced more hMCT8 mRNA and protein relative to the viral dose, which was present in various brain regions and localized to the cell membranes. Despite production of abundant hMCT8 mRNA and protein with ICV delivery, only IV delivered AAV9-hMCT8 targeted the choroid plexus and significantly increased brain T3 content and expression of Hairless.
Conclusions: These results indicate that MCT8 delivery to brain barriers by IV but not ICV injection is crucial for its proper function. MCT8 has no constitutive activity but acts through an increase in T3 entering the brain tissue. Increasing MCT8 expression in brain cell membranes, including neurons, is insufficient to produce an effect without an increase in brain T3 content. The correct hMCT8 isoform along with an optimized delivery method are critical for an effective gene therapy to provide functional MCT8 in the brain of patients with MCT8 mutations.
Conflict of interest statement
Author Disclosure Statement The authors have nothing to disclose.
Figures
Similar articles
-
AAV9-MCT8 Delivery at Juvenile Stage Ameliorates Neurological and Behavioral Deficits in a Mouse Model of MCT8-Deficiency.Thyroid. 2022 Jul;32(7):849-859. doi: 10.1089/thy.2022.0034. Epub 2022 May 20. Thyroid. 2022. PMID: 35350867 Free PMC article.
-
Mct8-deficient mice have increased energy expenditure and reduced fat mass that is abrogated by normalization of serum T3 levels.Endocrinology. 2013 Dec;154(12):4885-95. doi: 10.1210/en.2013-1150. Epub 2013 Sep 12. Endocrinology. 2013. PMID: 24029243 Free PMC article.
-
Tissue-specific thyroid hormone deprivation and excess in monocarboxylate transporter (mct) 8-deficient mice.Endocrinology. 2006 Sep;147(9):4036-43. doi: 10.1210/en.2006-0390. Epub 2006 May 18. Endocrinology. 2006. PMID: 16709608
-
Thyroid hormone transporters--functions and clinical implications.Nat Rev Endocrinol. 2015 Jul;11(7):406-17. doi: 10.1038/nrendo.2015.66. Epub 2015 May 5. Nat Rev Endocrinol. 2015. PMID: 25942657 Review.
-
Inherited defects of thyroid hormone-cell-membrane transport: review of recent findings.Curr Opin Endocrinol Diabetes Obes. 2013 Oct;20(5):434-40. doi: 10.1097/01.med.0000432531.03233.ad. Curr Opin Endocrinol Diabetes Obes. 2013. PMID: 23974772 Free PMC article. Review.
Cited by
-
Establishing Patient-Centered Outcomes for MCT8 Deficiency: Stakeholder Engagement and Systematic Literature Review.Neuropsychiatr Dis Treat. 2023 Oct 20;19:2195-2216. doi: 10.2147/NDT.S379703. eCollection 2023. Neuropsychiatr Dis Treat. 2023. PMID: 37881807 Free PMC article.
-
Thyroid Hormone Availability and Action during Brain Development in Rodents.Front Cell Neurosci. 2017 Aug 14;11:240. doi: 10.3389/fncel.2017.00240. eCollection 2017. Front Cell Neurosci. 2017. PMID: 28855863 Free PMC article.
-
Thyroid Hormone Supplementation Restores Cognitive Deficit, Insulin Signaling, and Neuroinflammation in the Hippocampus of a Sporadic Alzheimer's-like Disease Rat Model.Cells. 2024 Oct 30;13(21):1793. doi: 10.3390/cells13211793. Cells. 2024. PMID: 39513900 Free PMC article.
-
Monocarboxylate Transporter 8 Deficiency: From Pathophysiological Understanding to Therapy Development.Front Endocrinol (Lausanne). 2021 Sep 1;12:723750. doi: 10.3389/fendo.2021.723750. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 34539576 Free PMC article. Review.
-
Triac Treatment Prevents Neurodevelopmental and Locomotor Impairments in Thyroid Hormone Transporter Mct8/Oatp1c1 Deficient Mice.Int J Mol Sci. 2023 Feb 9;24(4):3452. doi: 10.3390/ijms24043452. Int J Mol Sci. 2023. PMID: 36834863 Free PMC article.
References
-
- Friesema EC, Ganguly S, Abdalla A, Manning Fox JE, Halestrap AP, Visser TJ. 2003. Identification of monocarboxylate transporter 8 as a specific thyroid hormone transporter. J Biol Chem 278:40128–40135 - PubMed
-
- Friesema EC, Grueters A, Biebermann H, Krude H, von Moers A, Reeser M, Barrett TG, Mancilla EE, Svensson J, Kester MH, Kuiper GG, Balkassmi S, Uitterlinden AG, Koehrle J, Rodien P, Halestrap AP, Visser TJ. 2004. Association between mutations in a thyroid hormone transporter and severe X-linked psychomotor retardation. Lancet 364:1435–1437 - PubMed
-
- Gika AD, Siddiqui A, Hulse AJ, Edwards S, Fallon P, McEntagart ME, Jan W, Josifova D, Lerman-Sagie T, Drummond J, Thompson E, Refetoff S, Bönnemann CG, Jungbluth H. 2010. White matter abnormalities and dystonic motor disorder associated with mutations in the SLC16A2 gene. Dev Med Child Neurol 52:475–482 - PMC - PubMed
-
- La Piana R, Vanasse M, Brais B, Bernard G. 2015. Myelination delay and Allan–Herndon–Dudley syndrome caused by a novel mutation in the SLC16A2 gene. J Child Neurol 30:1371–1374 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
