

Activation of the Yeast UBI4 Polyubiquitin Gene by Zap1 Transcription Factor via an Intragenic Promoter Is Critical for Zinc-deficient Growth
- PMID: 27432887
- PMCID: PMC5009263
- DOI: 10.1074/jbc.M116.743120
Activation of the Yeast UBI4 Polyubiquitin Gene by Zap1 Transcription Factor via an Intragenic Promoter Is Critical for Zinc-deficient Growth
Abstract
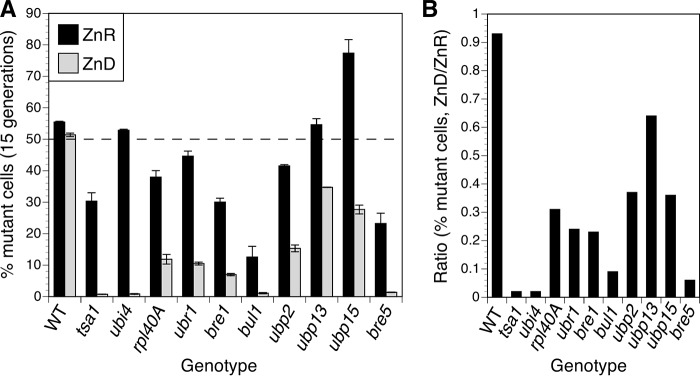
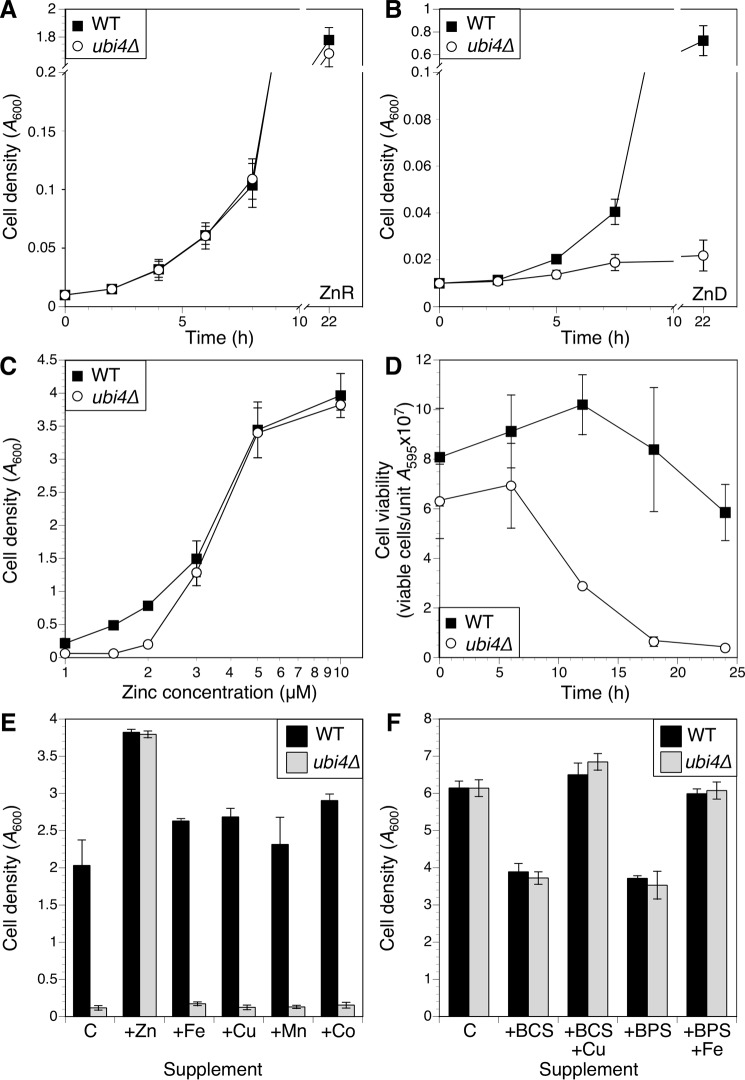
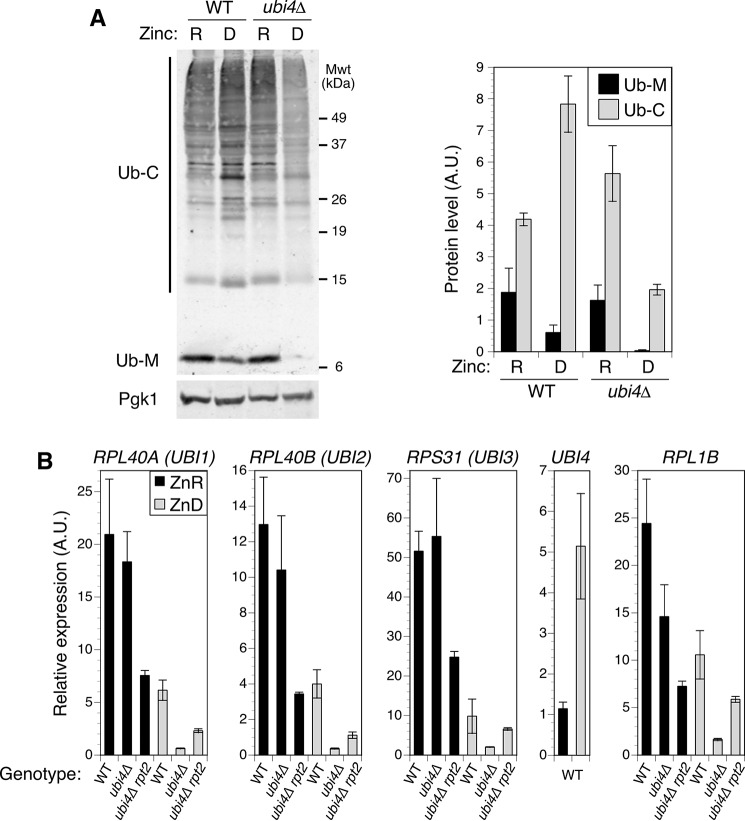
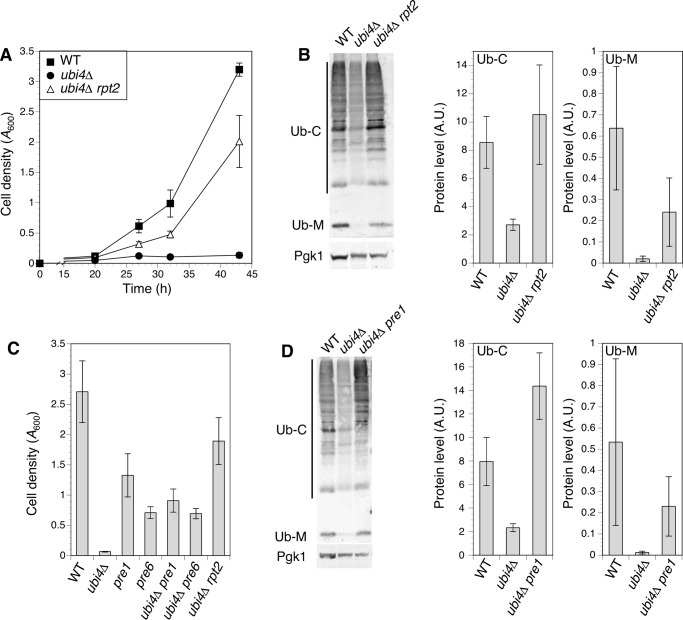
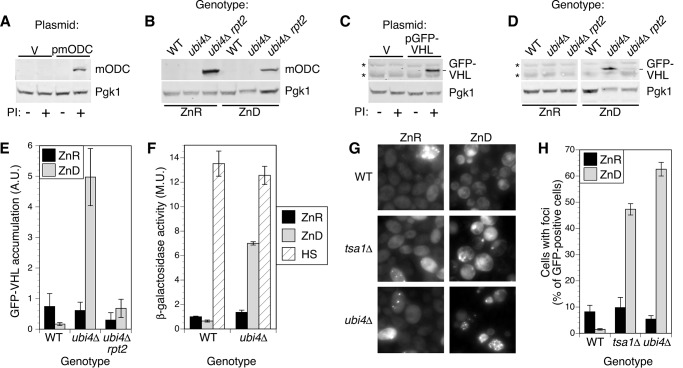
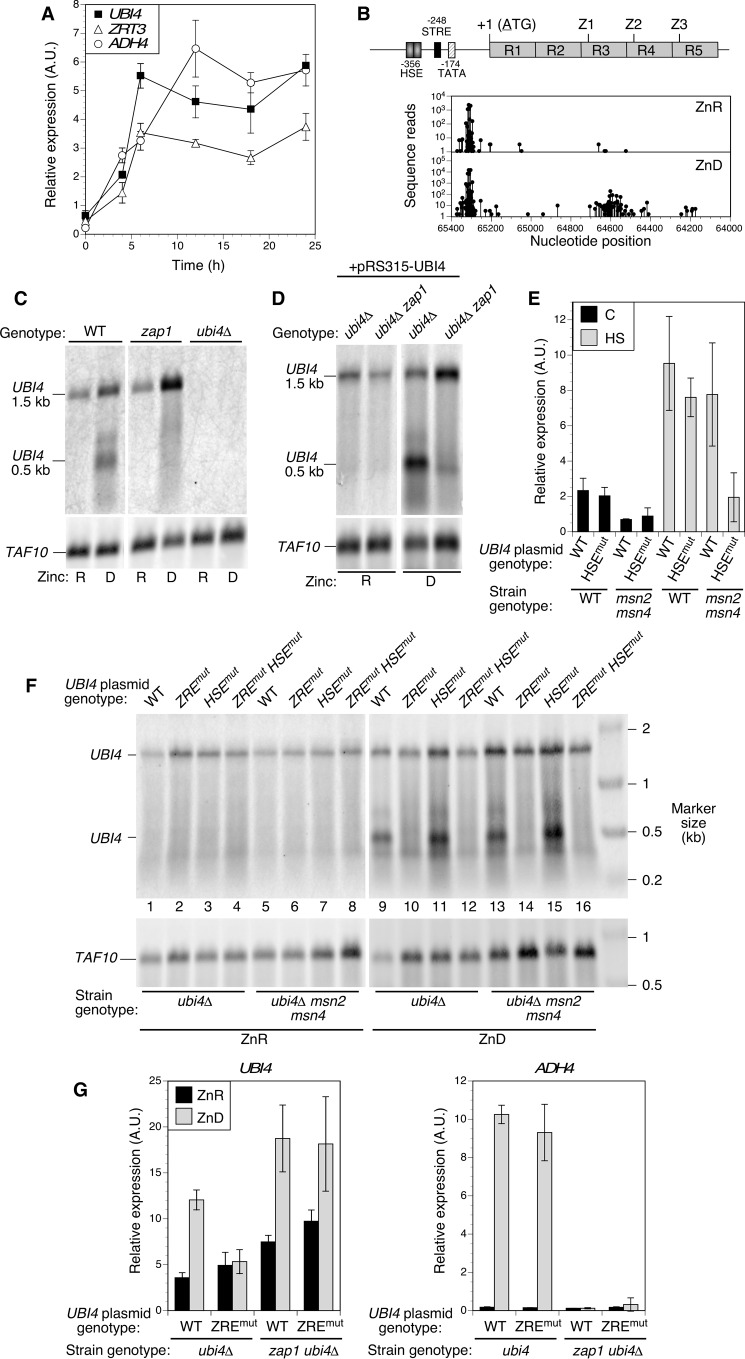
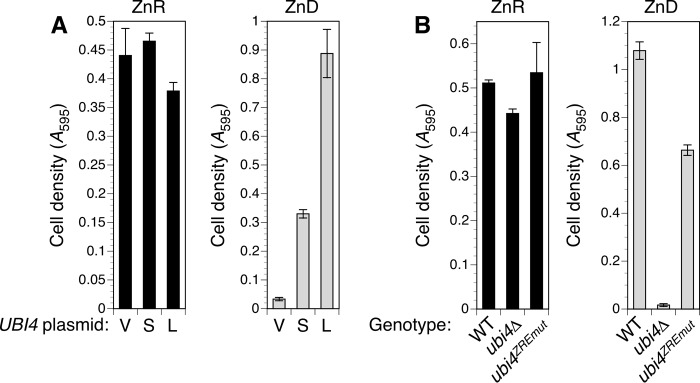
Stability of many proteins requires zinc. Zinc deficiency disrupts their folding, and the ubiquitin-proteasome system may help manage this stress. In Saccharomyces cerevisiae, UBI4 encodes five tandem ubiquitin monomers and is essential for growth in zinc-deficient conditions. Although UBI4 is only one of four ubiquitin-encoding genes in the genome, a dramatic decrease in ubiquitin was observed in zinc-deficient ubi4Δ cells. The three other ubiquitin genes were strongly repressed under these conditions, contributing to the decline in ubiquitin. In a screen for ubi4Δ suppressors, a hypomorphic allele of the RPT2 proteasome regulatory subunit gene (rpt2(E301K)) suppressed the ubi4Δ growth defect. The rpt2(E301K) mutation also increased ubiquitin accumulation in zinc-deficient cells, and by using a ubiquitin-independent proteasome substrate we found that proteasome activity was reduced. These results suggested that increased ubiquitin supply in suppressed ubi4Δ cells was a consequence of more efficient ubiquitin release and recycling during proteasome degradation. Degradation of a ubiquitin-dependent substrate was restored by the rpt2(E301K) mutation, indicating that ubiquitination is rate-limiting in this process. The UBI4 gene was induced ∼5-fold in low zinc and is regulated by the zinc-responsive Zap1 transcription factor. Surprisingly, Zap1 controls UBI4 by inducing transcription from an intragenic promoter, and the resulting truncated mRNA encodes only two of the five ubiquitin repeats. Expression of a short transcript alone complemented the ubi4Δ mutation, indicating that it is efficiently translated. Loss of Zap1-dependent UBI4 expression caused a growth defect in zinc-deficient conditions. Thus, the intragenic UBI4 promoter is critical to preventing ubiquitin deficiency in zinc-deficient cells.
Keywords: gene transcription; proteasome; transcription promoter; ubiquitin; yeast; zinc.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Zap1-dependent transcription from an alternative upstream promoter controls translation of RTC4 mRNA in zinc-deficient Saccharomyces cerevisiae.Mol Microbiol. 2017 Dec;106(5):678-689. doi: 10.1111/mmi.13851. Epub 2017 Oct 23. Mol Microbiol. 2017. PMID: 28963784 Free PMC article.
-
Yeast polyubiquitin gene UBI4 deficiency leads to early induction of apoptosis and shortened replicative lifespan.Cell Stress Chaperones. 2018 Jul;23(4):527-537. doi: 10.1007/s12192-017-0860-3. Epub 2017 Nov 7. Cell Stress Chaperones. 2018. PMID: 29116578 Free PMC article.
-
Roles of two activation domains in Zap1 in the response to zinc deficiency in Saccharomyces cerevisiae.J Biol Chem. 2011 Feb 25;286(8):6844-54. doi: 10.1074/jbc.M110.203927. Epub 2010 Dec 22. J Biol Chem. 2011. PMID: 21177862 Free PMC article.
-
Zinc sensing and regulation in yeast model systems.Arch Biochem Biophys. 2016 Dec 1;611:30-36. doi: 10.1016/j.abb.2016.02.031. Epub 2016 Mar 3. Arch Biochem Biophys. 2016. PMID: 26940262 Free PMC article. Review.
-
Homeostatic and adaptive responses to zinc deficiency in Saccharomyces cerevisiae.J Biol Chem. 2009 Jul 10;284(28):18565-9. doi: 10.1074/jbc.R900014200. Epub 2009 Apr 10. J Biol Chem. 2009. PMID: 19363031 Free PMC article. Review.
Cited by
-
Transcription factors and transporters in zinc homeostasis: lessons learned from fungi.Crit Rev Biochem Mol Biol. 2020 Feb;55(1):88-110. doi: 10.1080/10409238.2020.1742092. Epub 2020 Mar 19. Crit Rev Biochem Mol Biol. 2020. PMID: 32192376 Free PMC article. Review.
-
Near-infrared fluorescent northern blot.RNA. 2018 Dec;24(12):1871-1877. doi: 10.1261/rna.068213.118. Epub 2018 Sep 10. RNA. 2018. PMID: 30201850 Free PMC article.
-
The GIS2 Gene Is Repressed by a Zinc-Regulated Bicistronic RNA in Saccharomyces cerevisiae.Genes (Basel). 2018 Sep 19;9(9):462. doi: 10.3390/genes9090462. Genes (Basel). 2018. PMID: 30235899 Free PMC article.
-
An MSC2 Promoter-lacZ Fusion Gene Reveals Zinc-Responsive Changes in Sites of Transcription Initiation That Occur across the Yeast Genome.PLoS One. 2016 Sep 22;11(9):e0163256. doi: 10.1371/journal.pone.0163256. eCollection 2016. PLoS One. 2016. PMID: 27657924 Free PMC article.
-
The cellular economy of the Saccharomyces cerevisiae zinc proteome.Metallomics. 2018 Dec 12;10(12):1755-1776. doi: 10.1039/c8mt00269j. Metallomics. 2018. PMID: 30358795 Free PMC article.
References
-
- Vallee B. L., and Auld D. S. (1990) Zinc coordination, function, and structure of zinc enzymes and other proteins. Biochemistry 29, 5647–5659 - PubMed
-
- McCall K. A., Huang C., and Fierke C. A. (2000) Function and mechanism of zinc metalloenzymes. J. Nutr. 130, 1437S–1446S - PubMed
-
- Andreini C., Banci L., Bertini I., and Rosato A. (2006) Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 5, 196–201 - PubMed
-
- Andreini C., Bertini I., Cavallaro G., Holliday G. L., and Thornton J. M. (2009) Metal-MACiE: a database of metals involved in biological catalysis. Bioinformatics 25, 2088–2089 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
