

Biophysical characterization of the honeybee DSC1 orthologue reveals a novel voltage-dependent Ca2+ channel subfamily: CaV4
- PMID: 27432995
- PMCID: PMC4969797
- DOI: 10.1085/jgp.201611614
Biophysical characterization of the honeybee DSC1 orthologue reveals a novel voltage-dependent Ca2+ channel subfamily: CaV4
Abstract
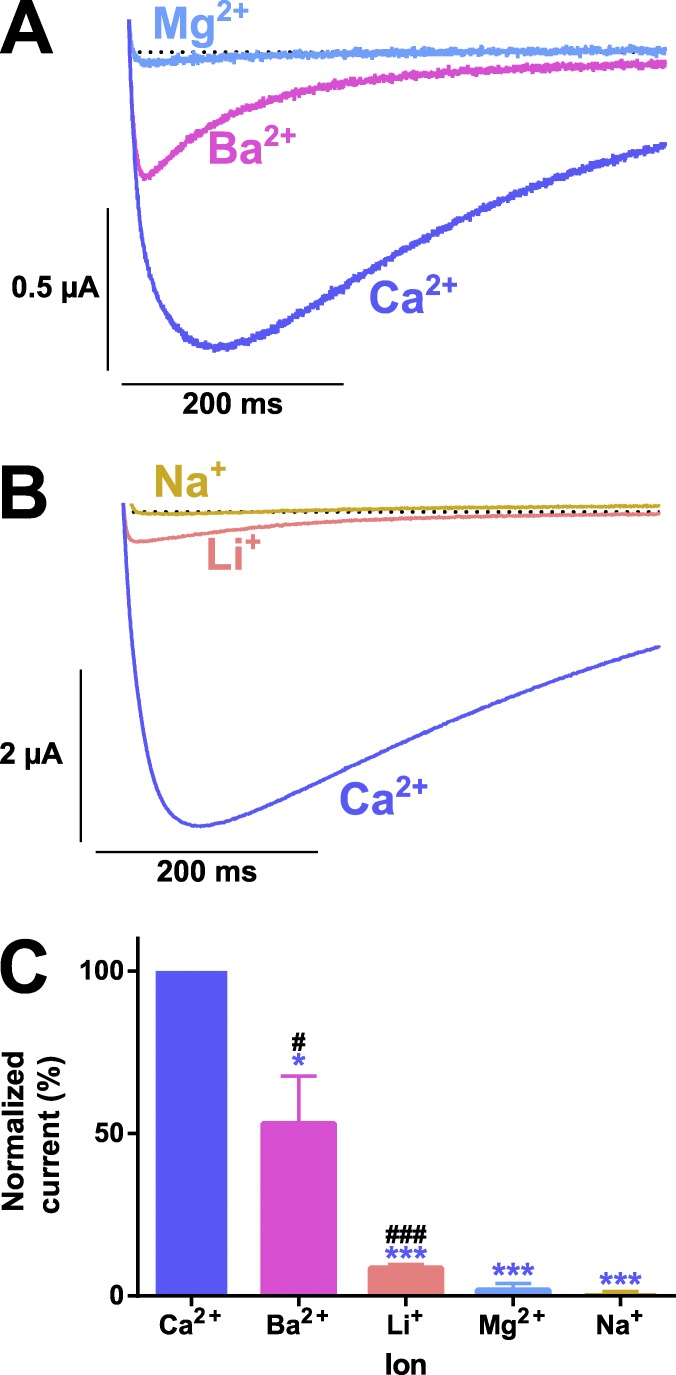
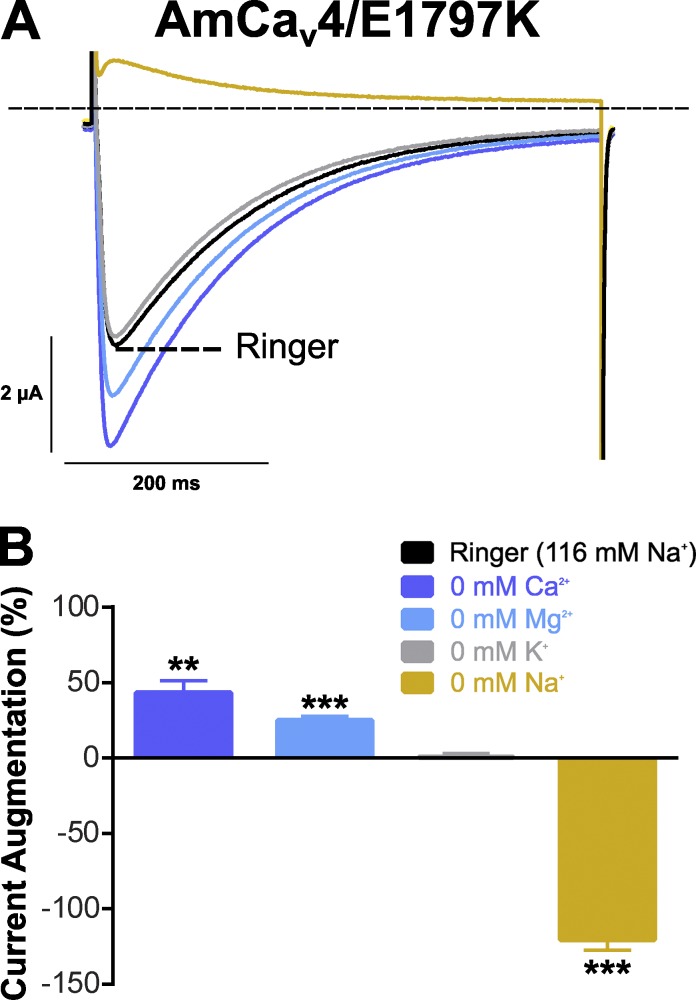
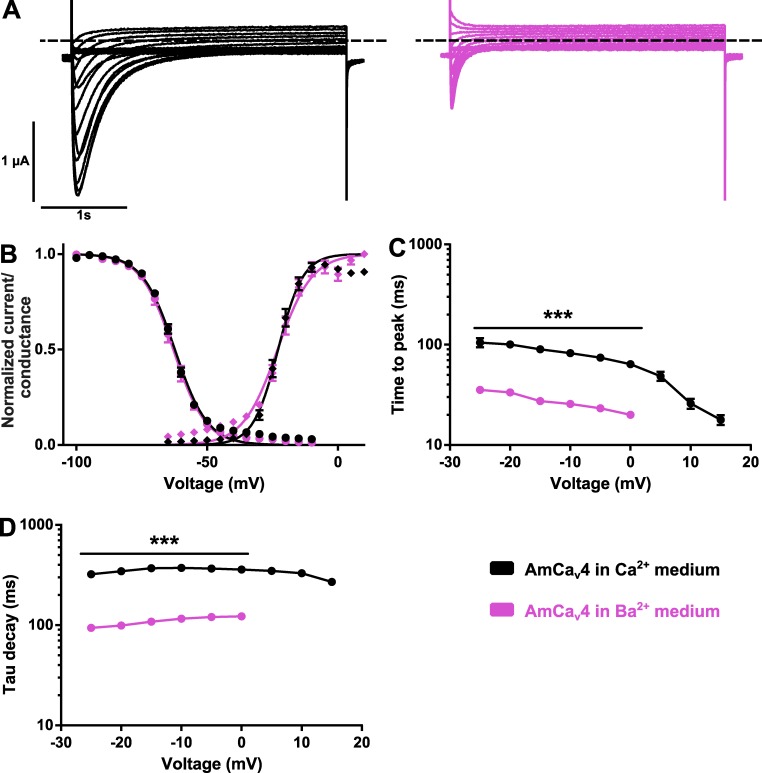
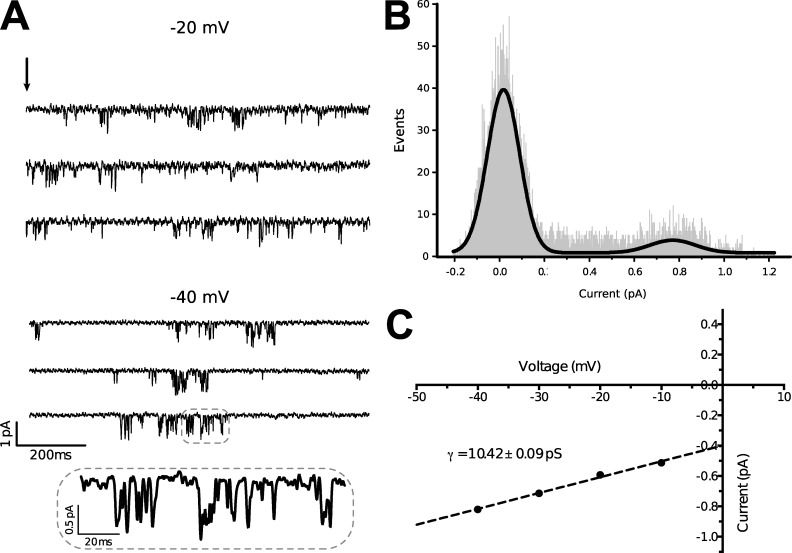
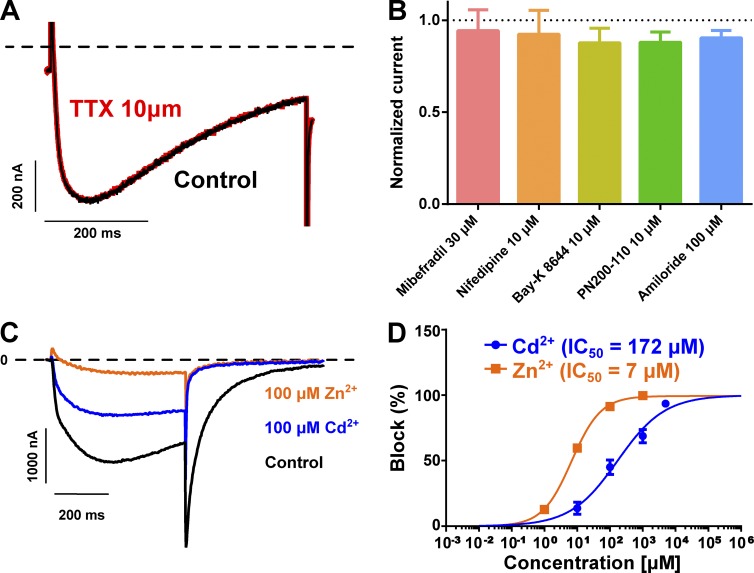
Bilaterian voltage-gated Na(+) channels (NaV) evolved from voltage-gated Ca(2+) channels (CaV). The Drosophila melanogaster Na(+) channel 1 (DSC1), which features a D-E-E-A selectivity filter sequence that is intermediate between CaV and NaV channels, is evidence of this evolution. Phylogenetic analysis has classified DSC1 as a Ca(2+)-permeable Na(+) channel belonging to the NaV2 family because of its sequence similarity with NaV channels. This is despite insect NaV2 channels (DSC1 and its orthologue in Blatella germanica, BSC1) being more permeable to Ca(2+) than Na(+) In this study, we report the cloning and molecular characterization of the honeybee (Apis mellifera) DSC1 orthologue. We reveal several sequence variations caused by alternative splicing, RNA editing, and genomic variations. Using the Xenopus oocyte heterologous expression system and the two-microelectrode voltage-clamp technique, we find that the channel exhibits slow activation and inactivation kinetics, insensitivity to tetrodotoxin, and block by Cd(2+) and Zn(2+) These characteristics are reminiscent of CaV channels. We also show a strong selectivity for Ca(2+) and Ba(2+) ions, marginal permeability to Li(+), and impermeability to Mg(2+) and Na(+) ions. Based on current ion channel nomenclature, the D-E-E-A selectivity filter, and the properties we have uncovered, we propose that DSC1 homologues should be classified as CaV4 rather than NaV2. Indeed, channels that contain the D-E-E-A selectivity sequence are likely to feature the same properties as the honeybee's channel, namely slow activation and inactivation kinetics and strong selectivity for Ca(2+) ions.
© 2016 Gosselin-Badaroudine et al.
Figures
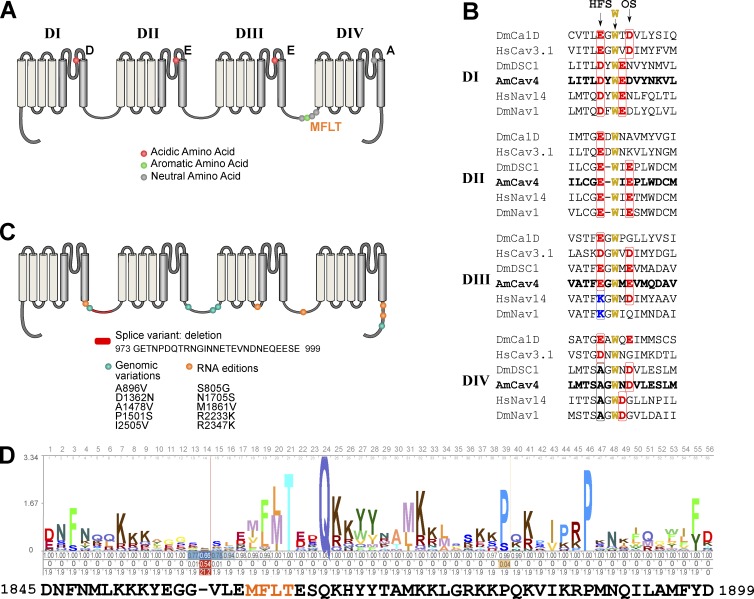
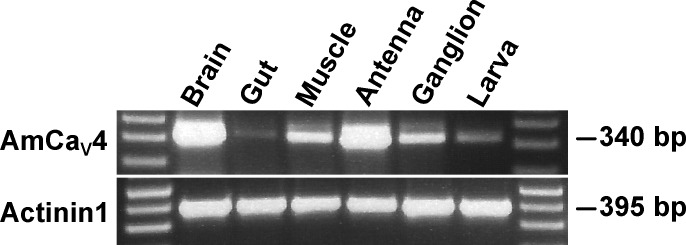
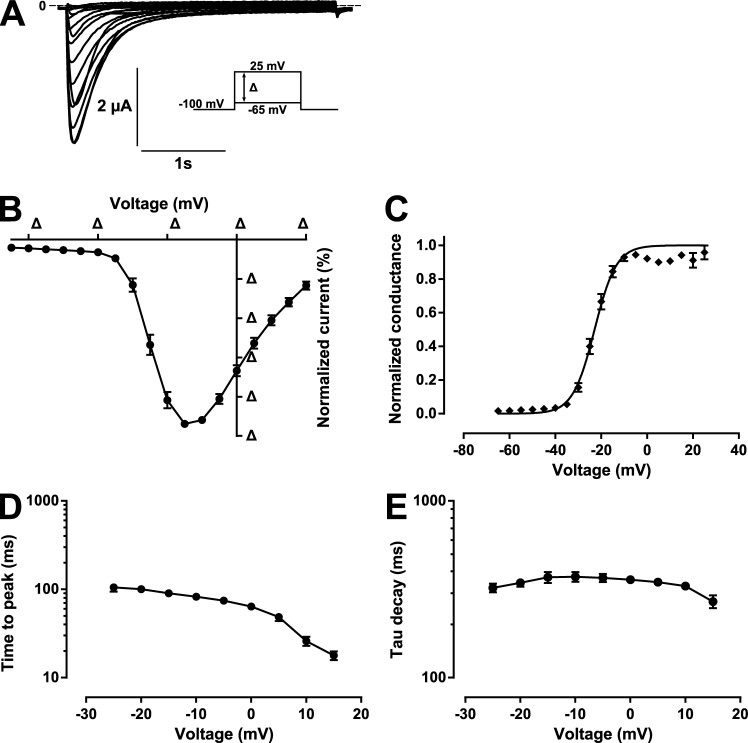
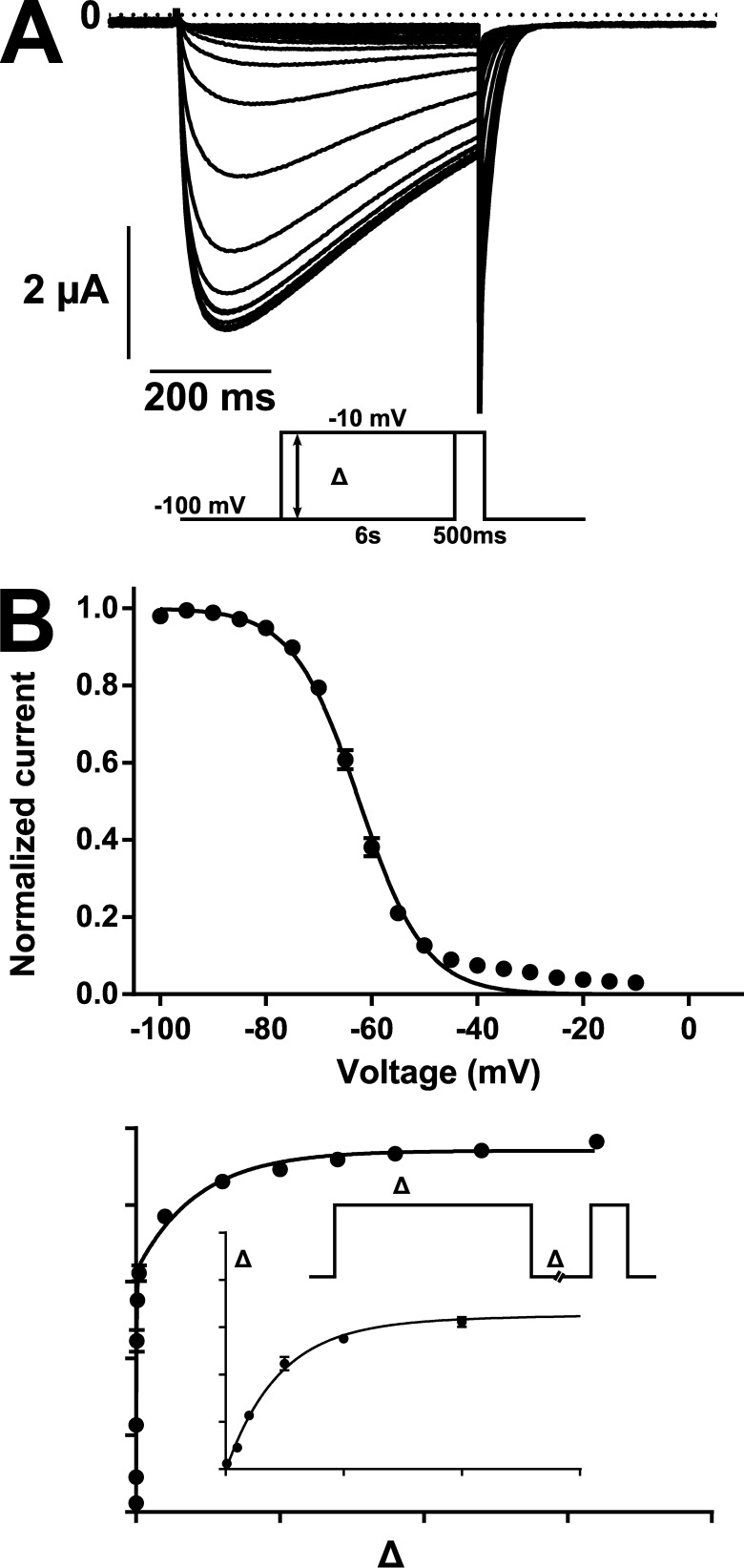

Comment in
-
What makes a sodium channel?J Gen Physiol. 2016 Aug;148(2):89-90. doi: 10.1085/jgp.201611652. Epub 2016 Jul 18. J Gen Physiol. 2016. PMID: 27432997 Free PMC article. No abstract available.
Similar articles
-
Honeybee CaV4 has distinct permeation, inactivation, and pharmacology from homologous NaV channels.J Gen Physiol. 2024 May 6;156(5):e202313509. doi: 10.1085/jgp.202313509. Epub 2024 Apr 1. J Gen Physiol. 2024. PMID: 38557788 Free PMC article.
-
Molecular characterization and functional expression of the DSC1 channel.Insect Biochem Mol Biol. 2011 Jul;41(7):451-8. doi: 10.1016/j.ibmb.2011.04.010. Epub 2011 May 7. Insect Biochem Mol Biol. 2011. PMID: 21571069 Free PMC article.
-
A voltage-gated calcium-selective channel encoded by a sodium channel-like gene.Neuron. 2004 Apr 8;42(1):101-12. doi: 10.1016/s0896-6273(04)00148-5. Neuron. 2004. PMID: 15066268 Free PMC article.
-
The Drosophila Sodium Channel 1 (DSC1): The founding member of a new family of voltage-gated cation channels.Pestic Biochem Physiol. 2015 May;120:36-9. doi: 10.1016/j.pestbp.2014.12.005. Epub 2014 Dec 9. Pestic Biochem Physiol. 2015. PMID: 25987218 Review.
-
Molecular pore structure of voltage-gated sodium and calcium channels.Braz J Med Biol Res. 1994 Dec;27(12):2781-802. Braz J Med Biol Res. 1994. PMID: 7550000 Review.
Cited by
-
Eukaryotic Voltage-Gated Sodium Channels: On Their Origins, Asymmetries, Losses, Diversification and Adaptations.Front Physiol. 2018 Nov 21;9:1406. doi: 10.3389/fphys.2018.01406. eCollection 2018. Front Physiol. 2018. PMID: 30519187 Free PMC article.
-
Honey bees long-lasting locomotor deficits after exposure to the diamide chlorantraniliprole are accompanied by brain and muscular calcium channels alterations.Sci Rep. 2019 Feb 15;9(1):2153. doi: 10.1038/s41598-019-39193-3. Sci Rep. 2019. PMID: 30770849 Free PMC article.
-
Honeybee locomotion is impaired by Am-CaV3 low voltage-activated Ca2+ channel antagonist.Sci Rep. 2017 Feb 1;7:41782. doi: 10.1038/srep41782. Sci Rep. 2017. PMID: 28145504 Free PMC article.
-
In silico identification and assessment of insecticide target sites in the genome of the small hive beetle, Aethina tumida.BMC Genomics. 2020 Feb 12;21(1):154. doi: 10.1186/s12864-020-6551-y. BMC Genomics. 2020. PMID: 32050907 Free PMC article.
-
Expression differences in Aphidius ervi (Hymenoptera: Braconidae) females reared on different aphid host species.PeerJ. 2017 Aug 21;5:e3640. doi: 10.7717/peerj.3640. eCollection 2017. PeerJ. 2017. PMID: 28852588 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
