

Anti-apoptotic Effects of Human Wharton's Jelly-derived Mesenchymal Stem Cells on Skeletal Muscle Cells Mediated via Secretion of XCL1
- PMID: 27434589
- PMCID: PMC5113102
- DOI: 10.1038/mt.2016.125
Anti-apoptotic Effects of Human Wharton's Jelly-derived Mesenchymal Stem Cells on Skeletal Muscle Cells Mediated via Secretion of XCL1
Abstract
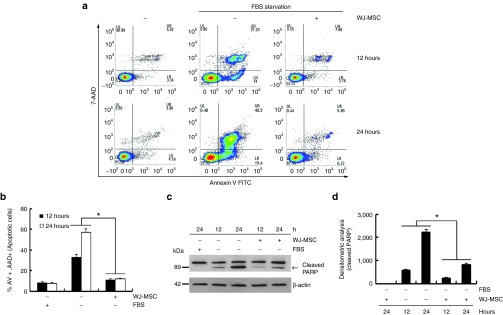
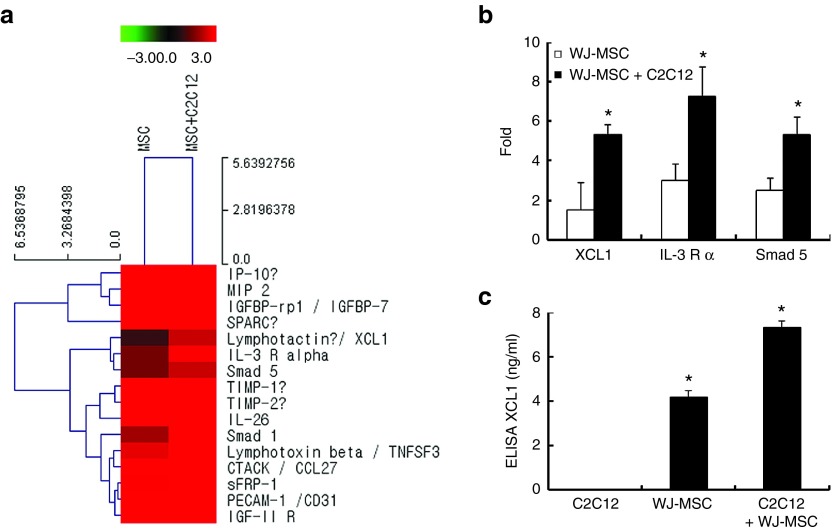
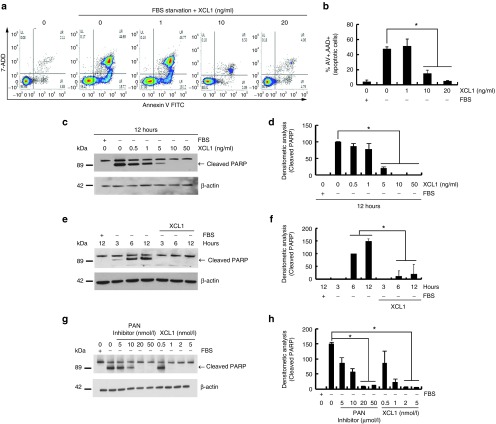
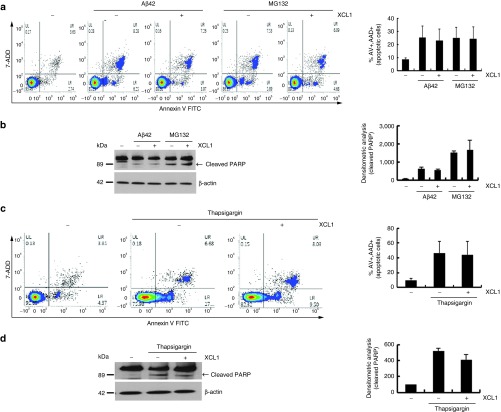
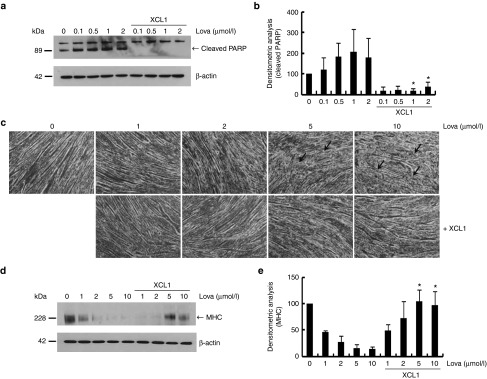
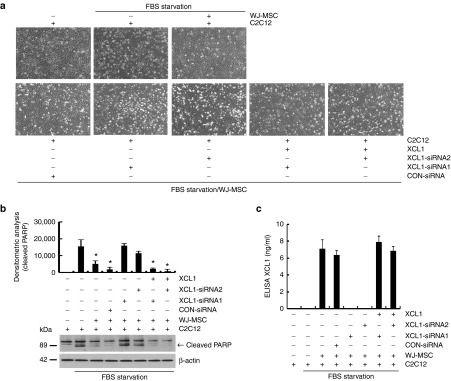
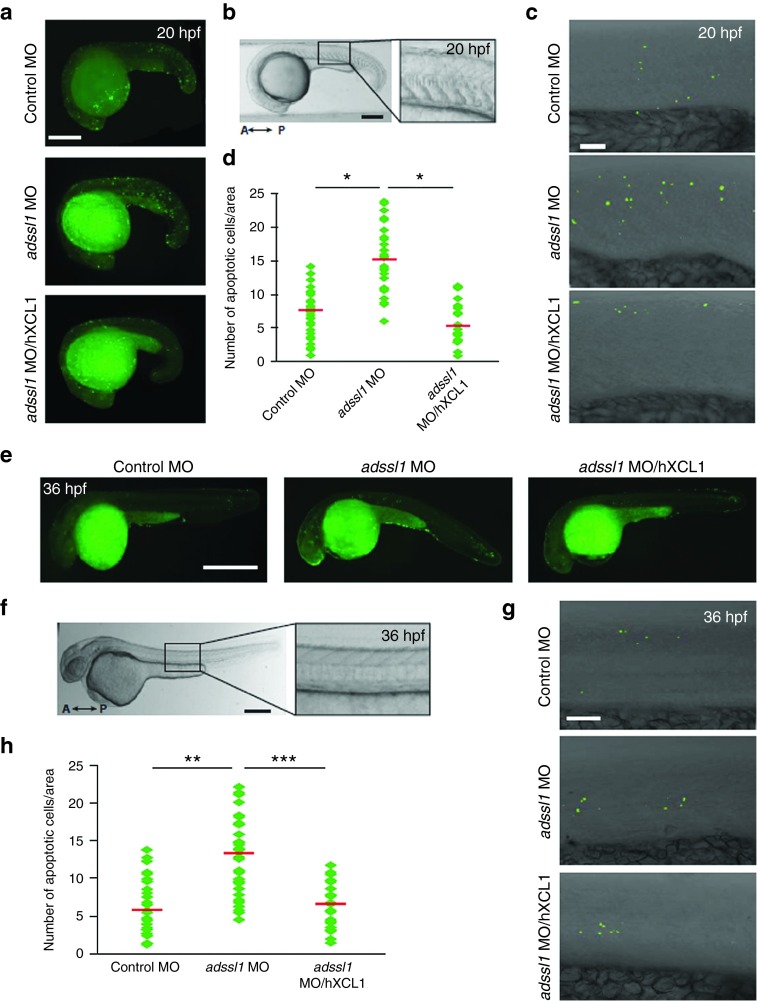
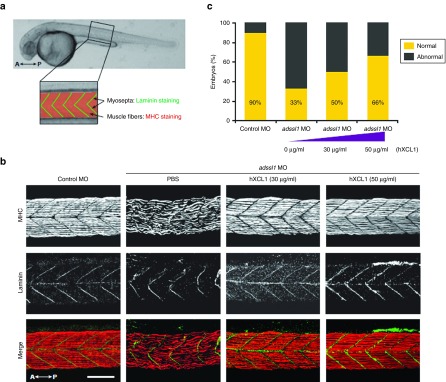
The role of Wharton's jelly-derived human mesenchymal stem cells (WJ-MSCs) in inhibiting muscle cell death has been elucidated in this study. Apoptosis induced by serum deprivation in mouse skeletal myoblast cell lines (C2C12) was significantly reduced when the cell lines were cocultured with WJ-MSCs. Antibody arrays indicated high levels of chemokine (C motif) ligand (XCL1) secretion by cocultured WJ-MSCs and XCL1 protein treatment resulted in complete inhibition of apoptosis in serum-starved C2C12 cells. Apoptosis of C2C12 cells and loss of differentiated C2C12 myotubes induced by lovastatin, another muscle cell death inducer, was also inhibited by XCL1 treatment. However, XCL1 treatment did not inhibit apoptosis of cell lines other than C2C12. When XCL1-siRNA pretreated WJ-MSCs were cocultured with serum-starved C2C12 cells, apoptosis was not inhibited, thus confirming that XCL1 is a key factor in preventing C2C12 cell apoptosis. We demonstrated the therapeutic effect of XCL1 on the zebrafish myopathy model, generated by knock down of a causative gene ADSSL1. Furthermore, the treatment of XCL1 resulted in significant recovery of the zebrafish skeletal muscle defects. These results suggest that human WJ-MSCs and XCL1 protein may act as promising and novel therapeutic agents for treatment of myopathies and other skeletal muscle diseases.
Figures
References
-
- Danial, NN and Korsmeyer, SJ (2004). Cell death: critical control points. Cell 116: 205–219. - PubMed
-
- Portt, L, Norman, G, Clapp, C, Greenwood, M and Greenwood, MT (2011). Anti-apoptosis and cell survival: a review. Biochim Biophys Acta 1813: 238–259. - PubMed
-
- Li, J and Yuan, J (2008). Caspases in apoptosis and beyond. Oncogene 27: 6194–6206. - PubMed
-
- Taylor, RC, Cullen, SP and Martin, SJ (2008). Apoptosis: controlled demolition at the cellular level. Nat Rev Mol Cell Biol 9: 231–241. - PubMed
-
- Adhihetty, PJ and Hood, DA (2003). Mechanisms of apoptosis in skeletal muscle. Basic Appl Myol 13: 171–179.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
