

Structural and thermodynamic basis of proline-induced transmembrane complex stabilization
- PMID: 27436065
- PMCID: PMC4951694
- DOI: 10.1038/srep29809
Structural and thermodynamic basis of proline-induced transmembrane complex stabilization
Abstract
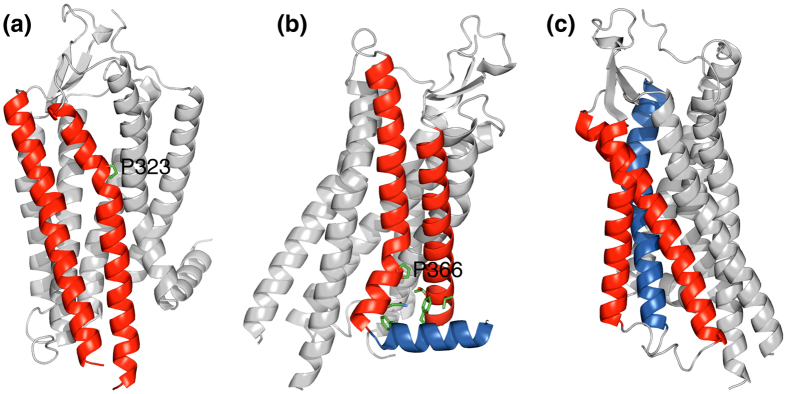
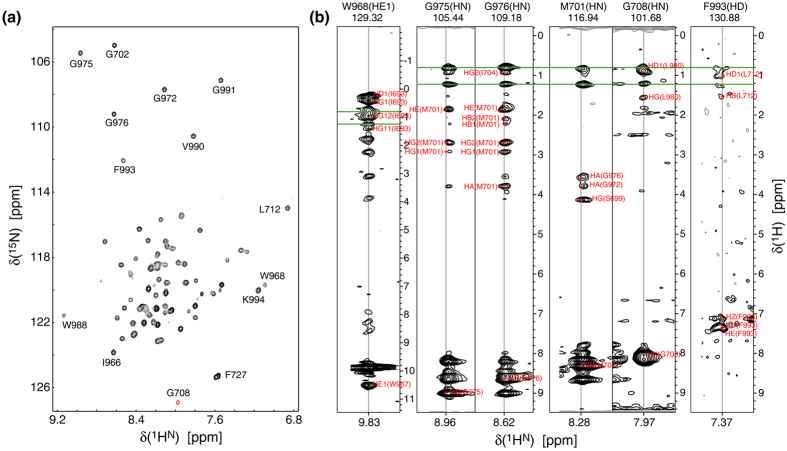
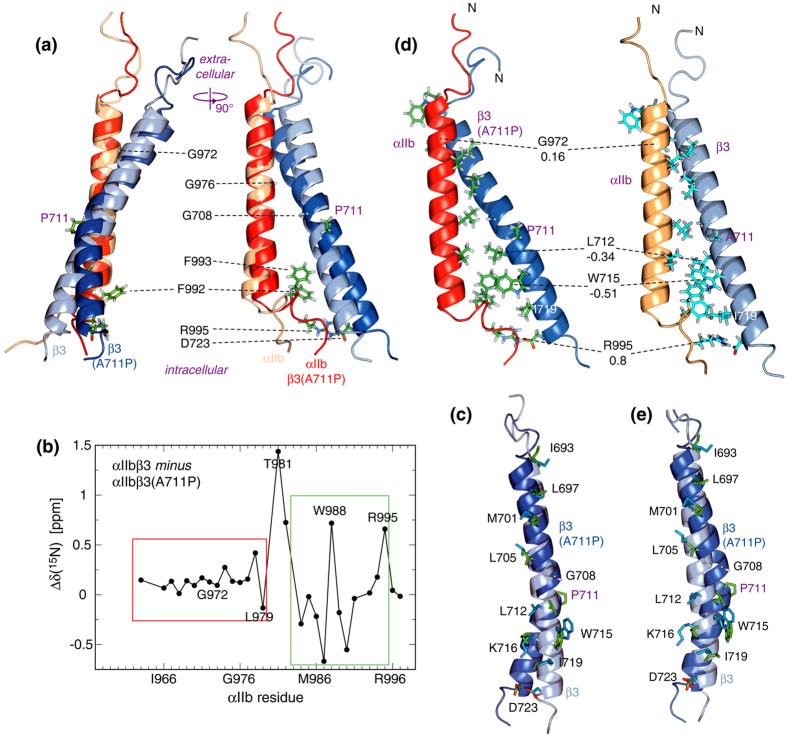
In membrane proteins, proline-mediated helix kinks are indispensable for the tight packing of transmembrane (TM) helices. However, kinks invariably affect numerous interhelical interactions, questioning the acceptance of proline substitutions and evolutionary origin of kinks. Here, we present the structural and thermodynamic basis of proline-induced integrin αIIbβ3 TM complex stabilization to understand the introduction of proline kinks in membrane proteins. In phospholipid bicelles, the A711P substitution in the center of the β3 TM helix changes the direction of adjacent helix segments to form a 35 ± 2° angle and predominantly repacks the segment in the inner membrane leaflet due to a swivel movement. This swivel repacks hydrophobic and electrostatic interhelical contacts within intracellular lipids, resulting in an overall TM complex stabilization of -0.82 ± 0.01 kcal/mol. Thus, proline substitutions can directly stabilize membrane proteins and such substitutions are proposed to follow the structural template of integrin αIIbβ3(A711P).
Figures
References
-
- Todd A. E., Orengo C. A. & Thornton J. M. Evolution of function in protein superfamilies, from a structural perspective. J Mol Biol 307, 1113–1143 (2001). - PubMed
-
- Schimmel P. R. & Flory P. J. Conformational Energies And Configurational Statistics Of Copolypeptides Containing L-Proline. J Mol Biol 34, 105–120 (1968). - PubMed
-
- Cordes F. S., Bright J. N. & Sansom M. S. P. Proline-induced distortions of transmembrane helices. J Mol Biol 323, 951–960 (2002). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
