

Rescue of perfluorooctanesulfonate (PFOS)-mediated Sertoli cell injury by overexpression of gap junction protein connexin 43
- PMID: 27436542
- PMCID: PMC4951654
- DOI: 10.1038/srep29667
Rescue of perfluorooctanesulfonate (PFOS)-mediated Sertoli cell injury by overexpression of gap junction protein connexin 43
Abstract
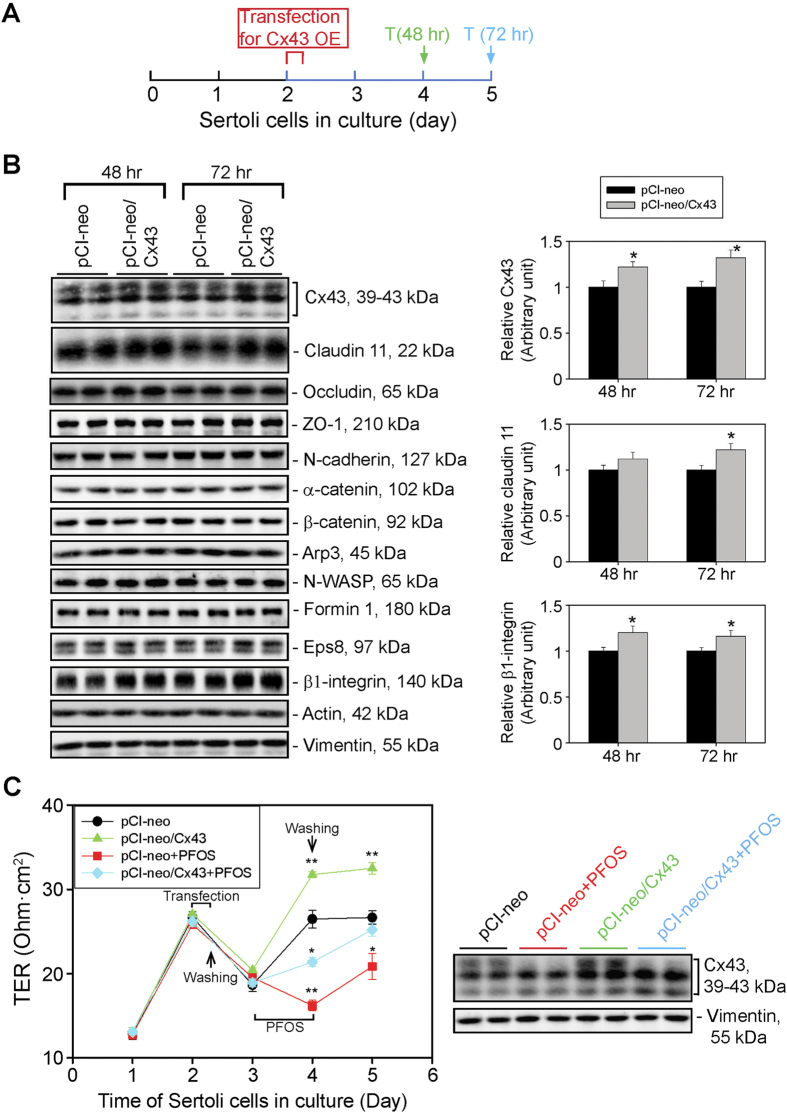
Perfluorooctanesulfonate (PFOS) is an environmental toxicant used in developing countries, including China, as a stain repellent for clothing, carpets and draperies, but it has been banned in the U.S. and Canada since the late 2000s. PFOS perturbed the Sertoli cell tight junction (TJ)-permeability barrier, causing disruption of actin microfilaments in cell cytosol, perturbing the localization of cell junction proteins (e.g., occluden-ZO-1, N-cadherin-ß-catenin). These changes destabilized Sertoli cell blood-testis barrier (BTB) integrity. These findings suggest that human exposure to PFOS might induce BTB dysfunction and infertility. Interestingly, PFOS-induced Sertoli cell injury associated with a down-regulation of the gap junction (GJ) protein connexin43 (Cx43). We next investigated if overexpression of Cx43 in Sertoli cells could rescue the PFOS-induced cell injury. Indeed, overexpression of Cx43 in Sertoli cells with an established TJ-barrier blocked the disruption in PFOS-induced GJ-intercellular communication, resulting in the re-organization of actin microfilaments, which rendered them similar to those in control cells. Furthermore, cell adhesion proteins that utilized F-actin for attachment became properly distributed at the cell-cell interface, resealing the disrupted TJ-barrier. In summary, Cx43 is a good target that might be used to manage PFOS-induced reproductive dysfunction.
Figures




References
-
- Balse E. et al.. Dynamic of ion channel expression at the plasmam membrane of cardiomyocytes. Physiol Rev 92, 1317–1358 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
