

CD47-blocking antibodies restore phagocytosis and prevent atherosclerosis
- PMID: 27437576
- PMCID: PMC4980260
- DOI: 10.1038/nature18935
CD47-blocking antibodies restore phagocytosis and prevent atherosclerosis
Abstract
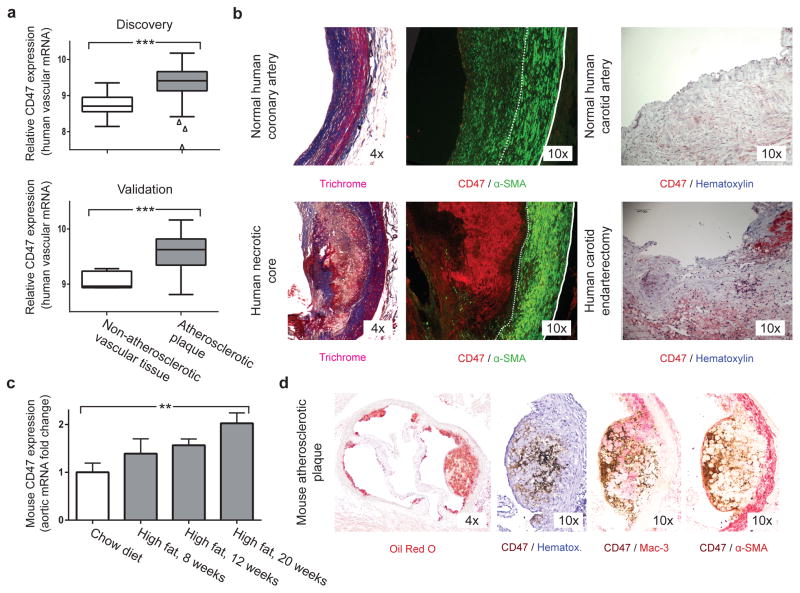
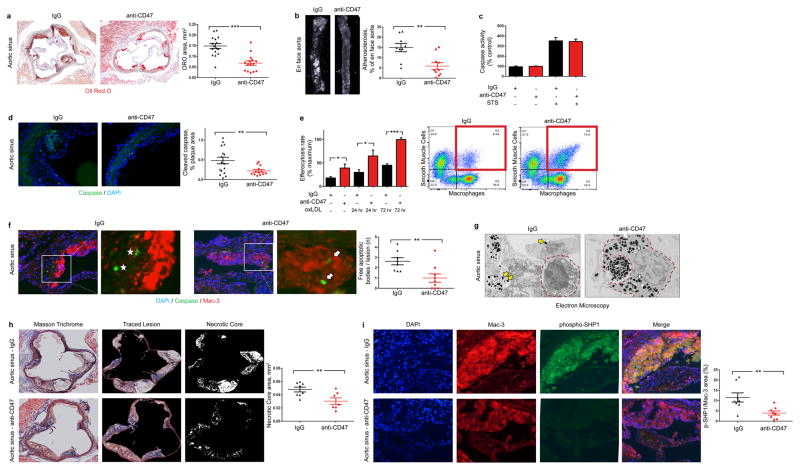
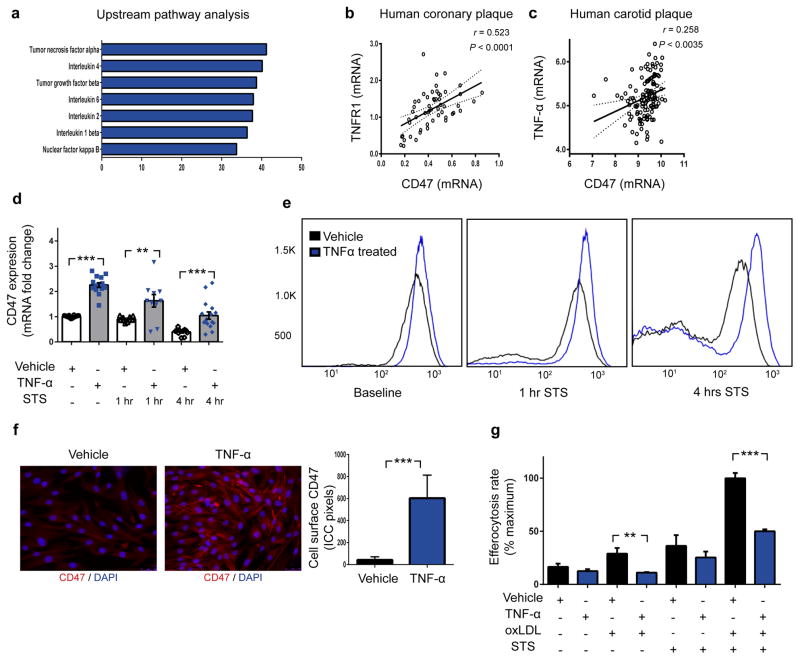
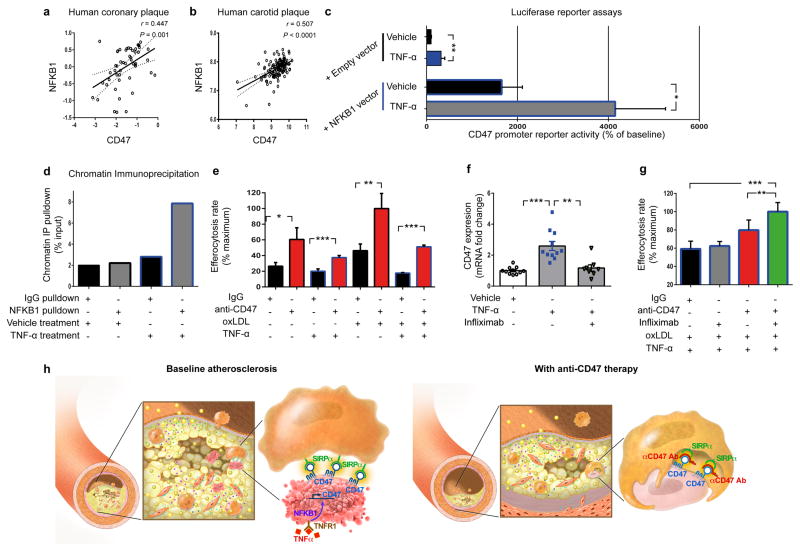
Atherosclerosis is the disease process that underlies heart attack and stroke. Advanced lesions at risk of rupture are characterized by the pathological accumulation of diseased vascular cells and apoptotic cellular debris. Why these cells are not cleared remains unknown. Here we show that atherogenesis is associated with upregulation of CD47, a key anti-phagocytic molecule that is known to render malignant cells resistant to programmed cell removal, or 'efferocytosis'. We find that administration of CD47-blocking antibodies reverses this defect in efferocytosis, normalizes the clearance of diseased vascular tissue, and ameliorates atherosclerosis in multiple mouse models. Mechanistic studies implicate the pro-atherosclerotic factor TNF-α as a fundamental driver of impaired programmed cell removal, explaining why this process is compromised in vascular disease. Similar to recent observations in cancer, impaired efferocytosis appears to play a pathogenic role in cardiovascular disease, but is not a fixed defect and may represent a novel therapeutic target.
Conflict of interest statement
The authors declare competing financial interests: N.J.L. and I.L.W. have filed a patent describing inhibition of CD47 as a method to prevent atherosclerosis.
Figures

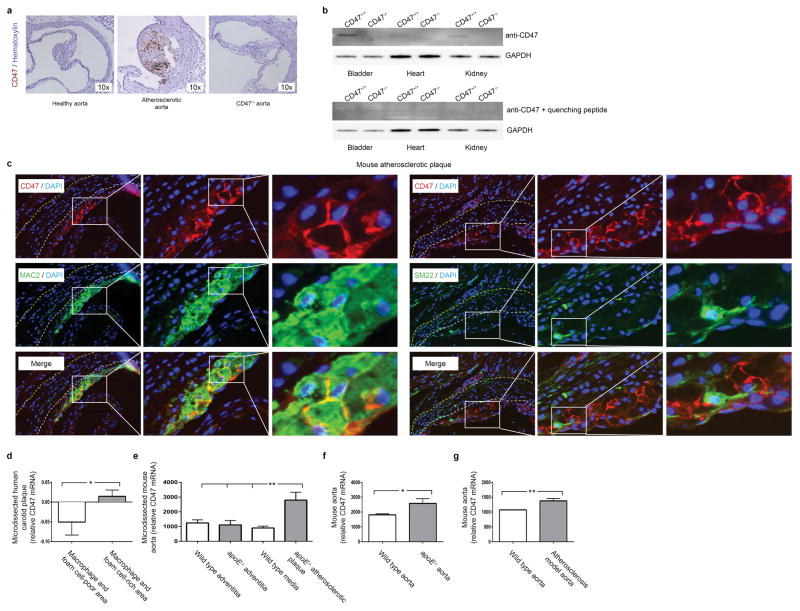
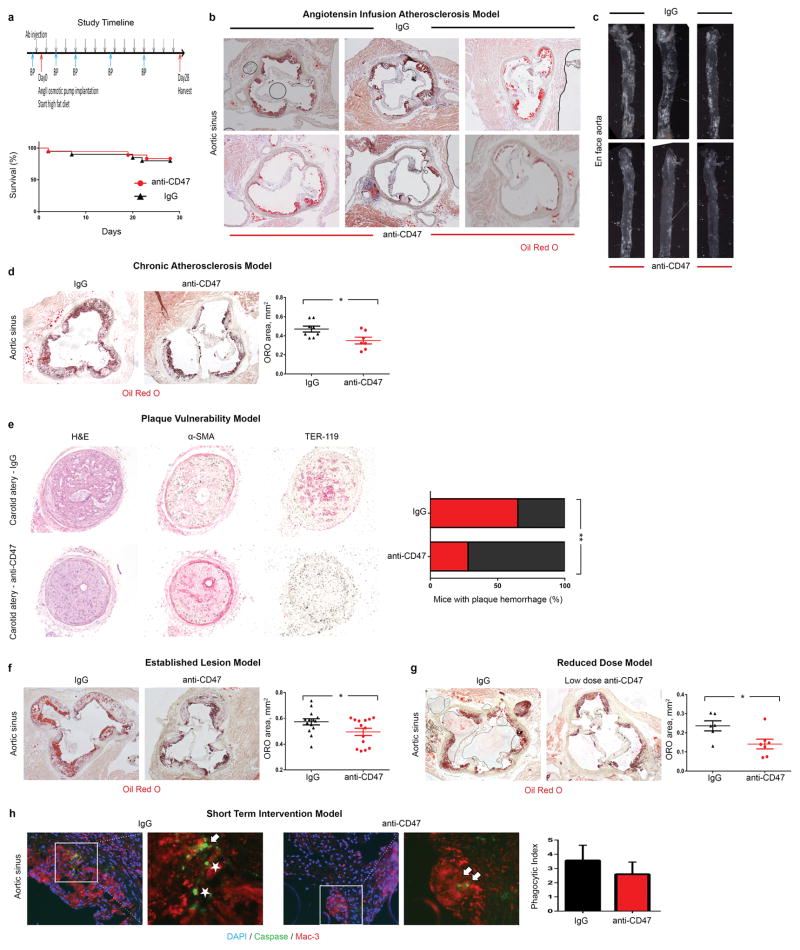
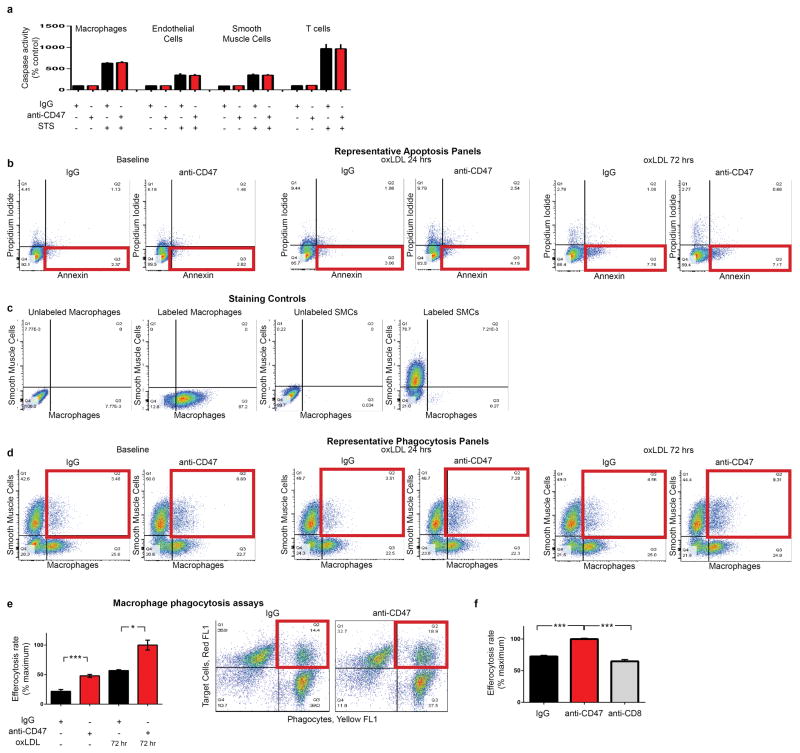
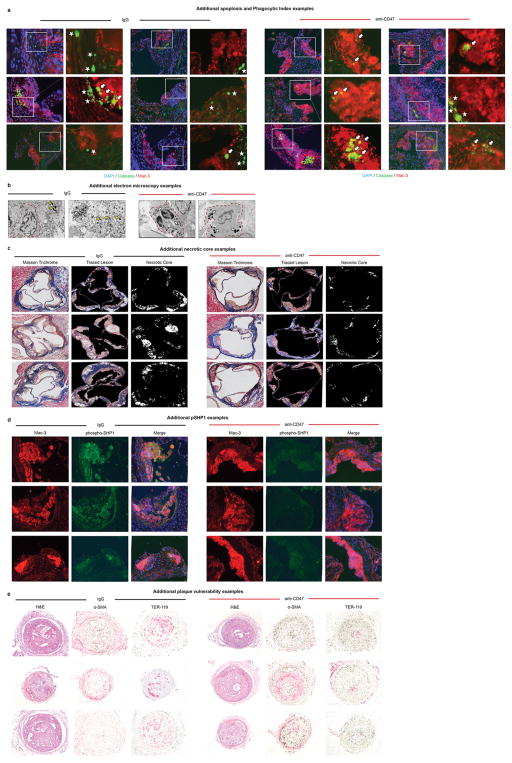
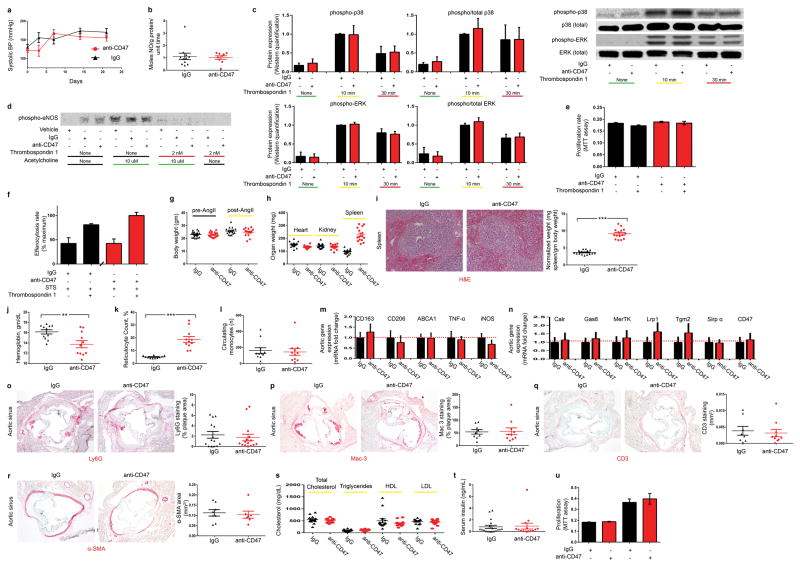
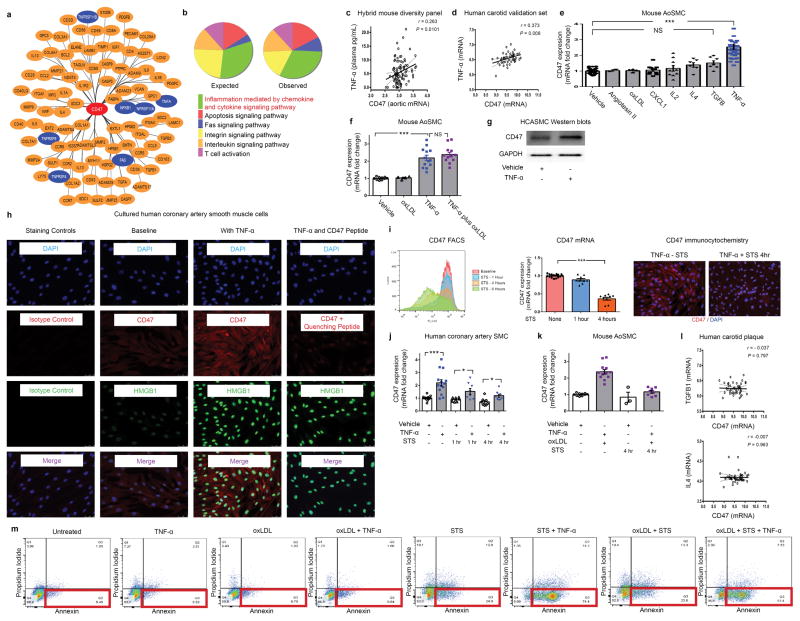
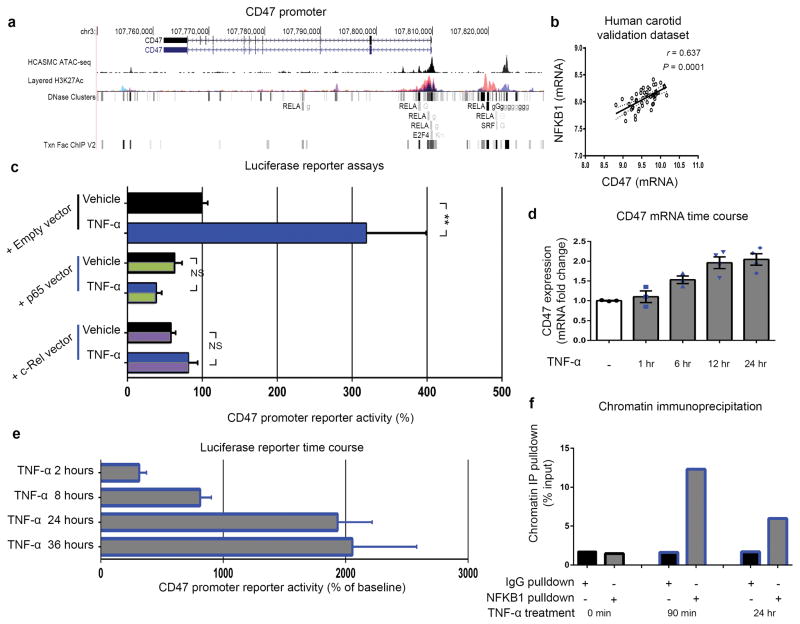
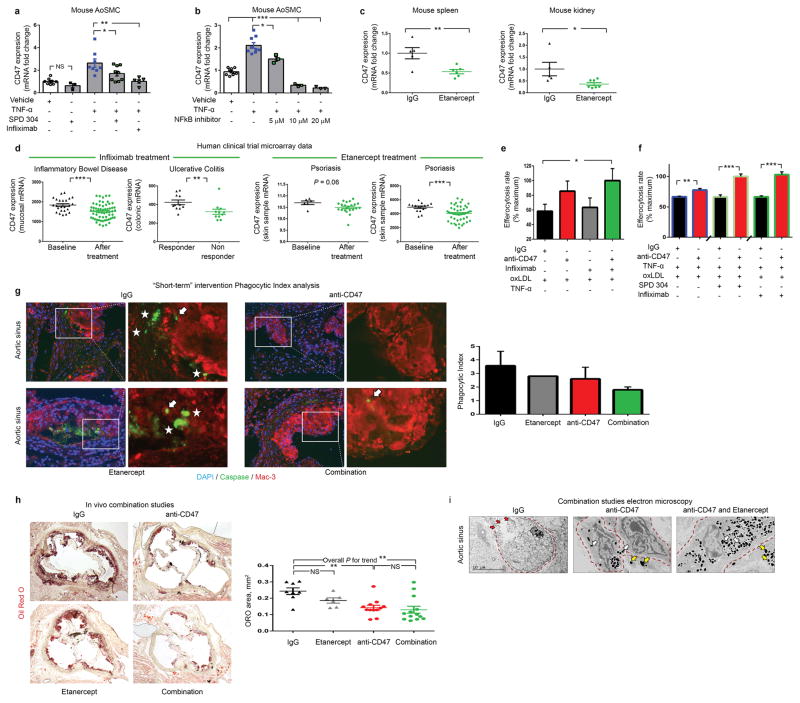
Comment in
-
Heart disease: Death-defying plaque cells.Nature. 2016 Aug 4;536(7614):32-3. doi: 10.1038/nature18916. Epub 2016 Jul 20. Nature. 2016. PMID: 27437578 Free PMC article.
-
Atherosclerosis: Treating atherosclerosis with antitumour antibodies.Nat Rev Cardiol. 2016 Sep;13(9):507. doi: 10.1038/nrcardio.2016.125. Epub 2016 Aug 4. Nat Rev Cardiol. 2016. PMID: 27489189 No abstract available.
References
-
- Libby P, Ridker PM, Hansson GK. Progress and challenges in translating the biology of atherosclerosis. Nature. 2011;473:317–25. - PubMed
-
- Schrijvers DM, De Meyer GR, Kockx MM, Herman AG, Martinet W. Phagocytosis of apoptotic cells by macrophages is impaired in atherosclerosis. Arterioscler Thromb Vasc Biol. 2005;25:1256–61. - PubMed
-
- Oldenborg PA, Zheleznyak A, Fang YF, Lagenaur CF, Gresham HD, Lindberg FP. Role of CD47 as a marker of self on red blood cells. Science. 2000;288:2051–4. - PubMed
-
- Chao MP, Majeti R, Weissman IL. Programmed cell removal: a new obstacle in the road to developing cancer. Nat Rev Cancer. 2012;12:58–67. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
