

ASPP2 Is a Novel Pan-Ras Nanocluster Scaffold
- PMID: 27437940
- PMCID: PMC4954646
- DOI: 10.1371/journal.pone.0159677
ASPP2 Is a Novel Pan-Ras Nanocluster Scaffold
Abstract
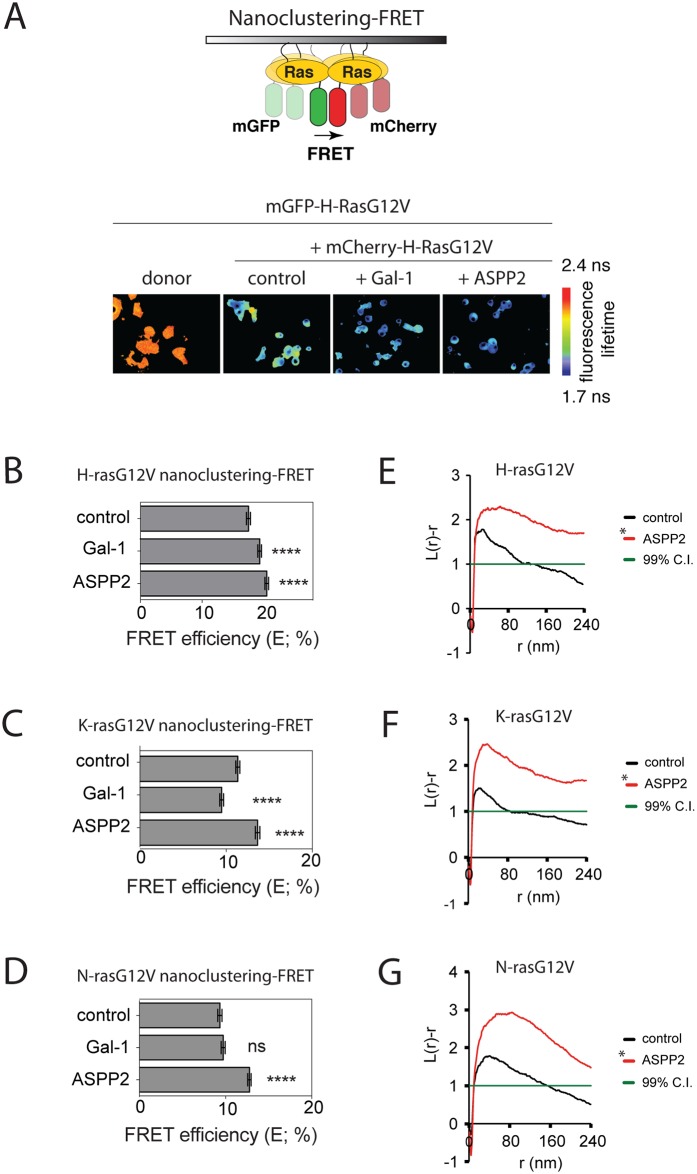
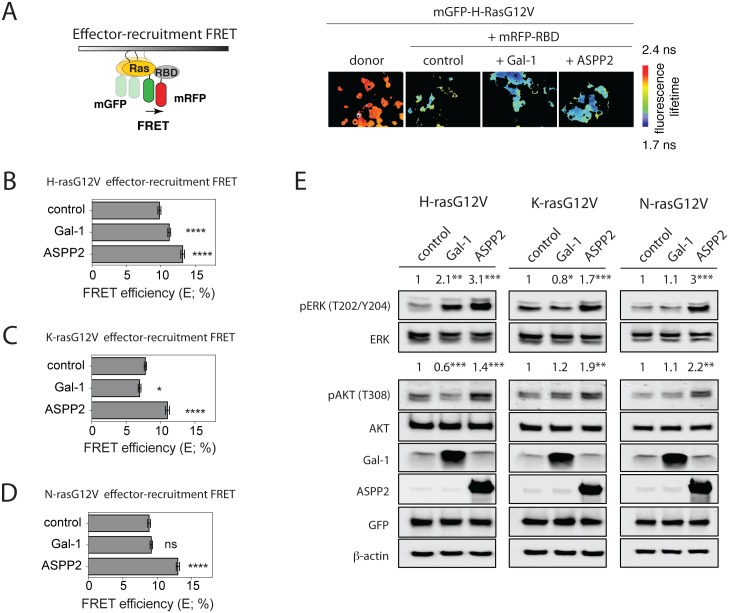
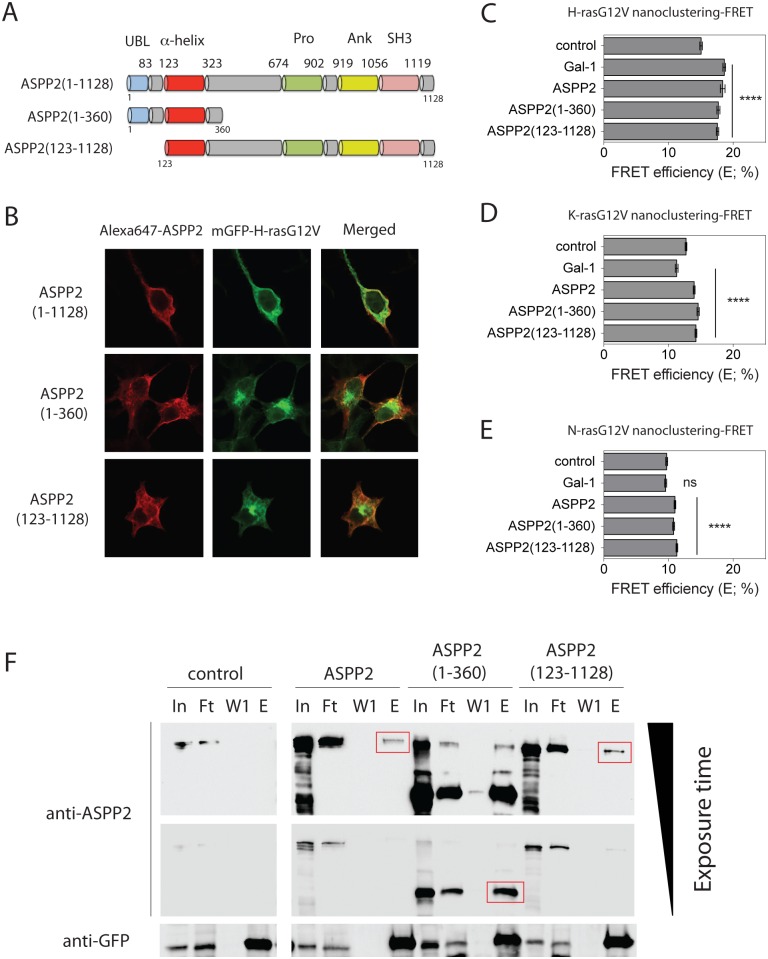
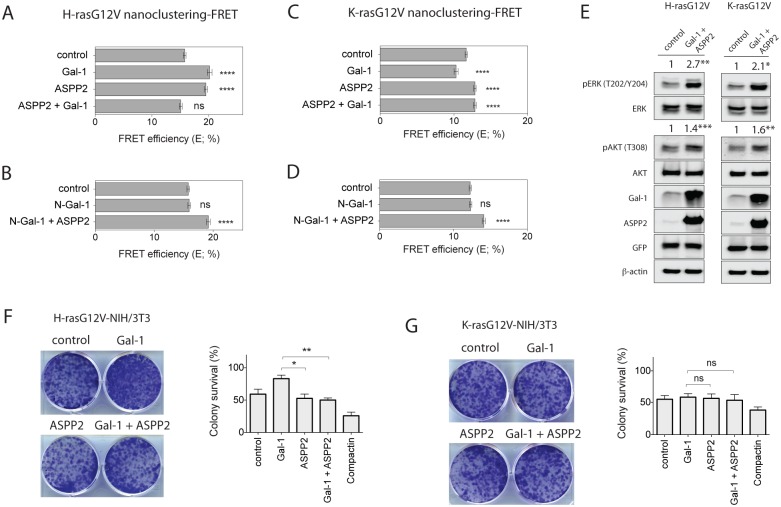
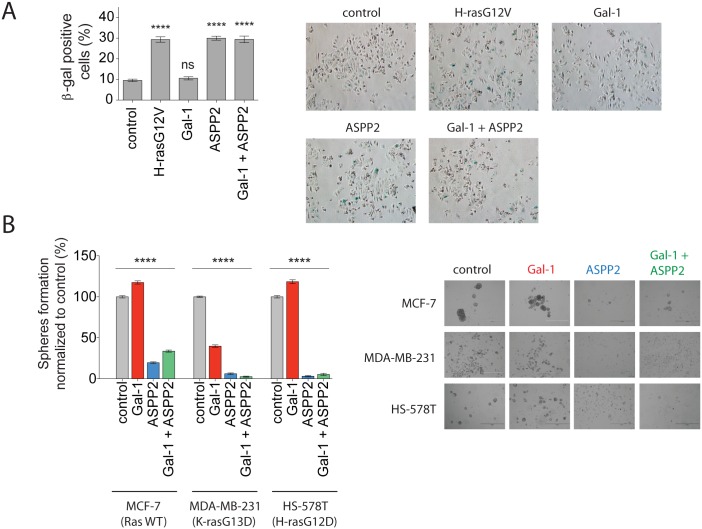
Ras-induced senescence mediated through ASPP2 represents a barrier to tumour formation. It is initiated by ASPP2's interaction with Ras at the plasma membrane, which stimulates the Raf/MEK/ERK signaling cascade. Ras to Raf signalling requires Ras to be organized in nanoscale signalling complexes, called nanocluster. We therefore wanted to investigate whether ASPP2 affects Ras nanoclustering. Here we show that ASPP2 increases the nanoscale clustering of all oncogenic Ras isoforms, H-ras, K-ras and N-ras. Structure-function analysis with ASPP2 truncation mutants suggests that the nanocluster scaffolding activity of ASPP2 converges on its α-helical domain. While ASPP2 increased effector recruitment and stimulated ERK and AKT phosphorylation, it did not increase colony formation of RasG12V transformed NIH/3T3 cells. By contrast, ASPP2 was able to suppress the transformation enhancing ability of the nanocluster scaffold Gal-1, by competing with the specific effect of Gal-1 on H-rasG12V- and K-rasG12V-nanoclustering, thus imposing ASPP2's ERK and AKT signalling signature. Similarly, ASPP2 robustly induced senescence and strongly abrogated mammosphere formation irrespective of whether it was expressed alone or together with Gal-1, which by itself showed the opposite effect in Ras wt or H-ras mutant breast cancer cells. Our results suggest that Gal-1 and ASPP2 functionally compete in nanocluster for active Ras on the plasma membrane. ASPP2 dominates the biological outcome, thus switching from a Gal-1 supported growth-promoting setting to a senescence inducing and stemness suppressive program in cancer cells. Our results support Ras nanocluster as major integrators of tumour fate decision events.
Conflict of interest statement
Figures
References
-
- Serrano M, Lin AW, McCurrach ME, Beach D, Lowe SW. Oncogenic ras provokes premature cell senescence associated with accumulation of p53 and p16INK4a. Cell. 1997;88: 593–602. - PubMed
-
- Yan J, Roy S, Apolloni A, Lane A, Hancock JF. Ras isoforms vary in their ability to activate Raf-1 and phosphoinositide 3-kinase. J Biol Chem. 1998;273: 24052–24056. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
