

Functional characterization of the ribosome biogenesis factors PES, BOP1, and WDR12 (PeBoW), and mechanisms of defective cell growth and proliferation caused by PeBoW deficiency in Arabidopsis
- PMID: 27440937
- PMCID: PMC5014167
- DOI: 10.1093/jxb/erw288
Functional characterization of the ribosome biogenesis factors PES, BOP1, and WDR12 (PeBoW), and mechanisms of defective cell growth and proliferation caused by PeBoW deficiency in Arabidopsis
Abstract
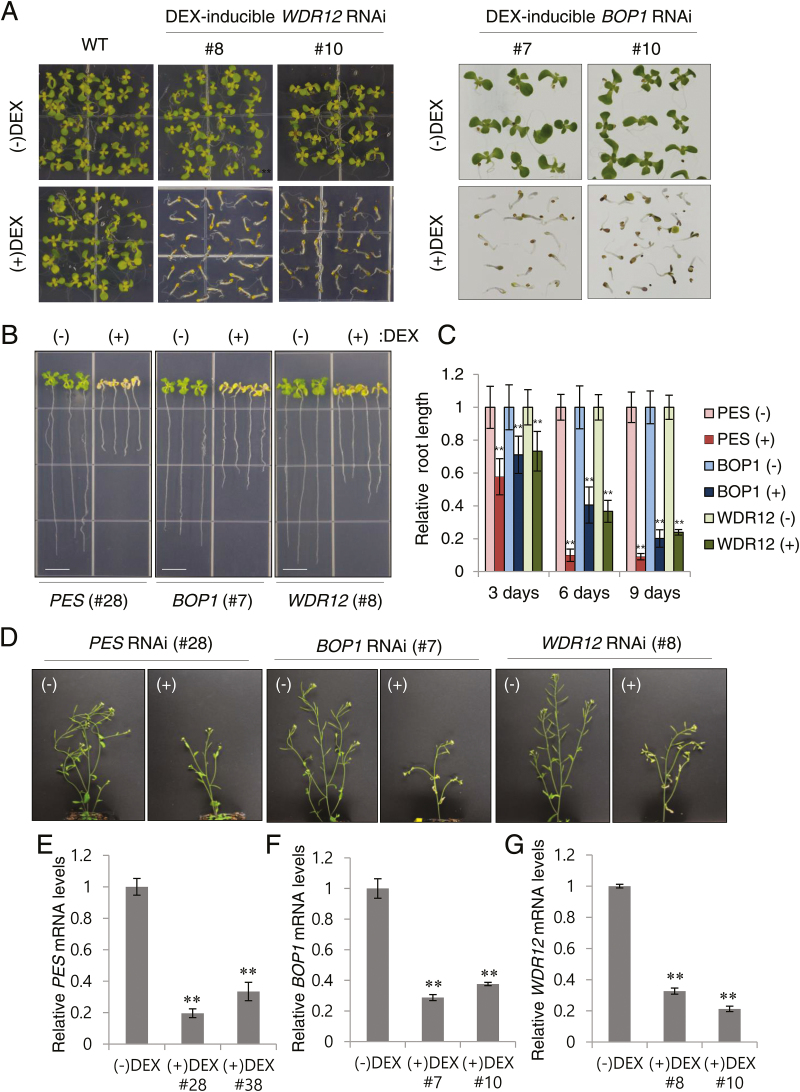
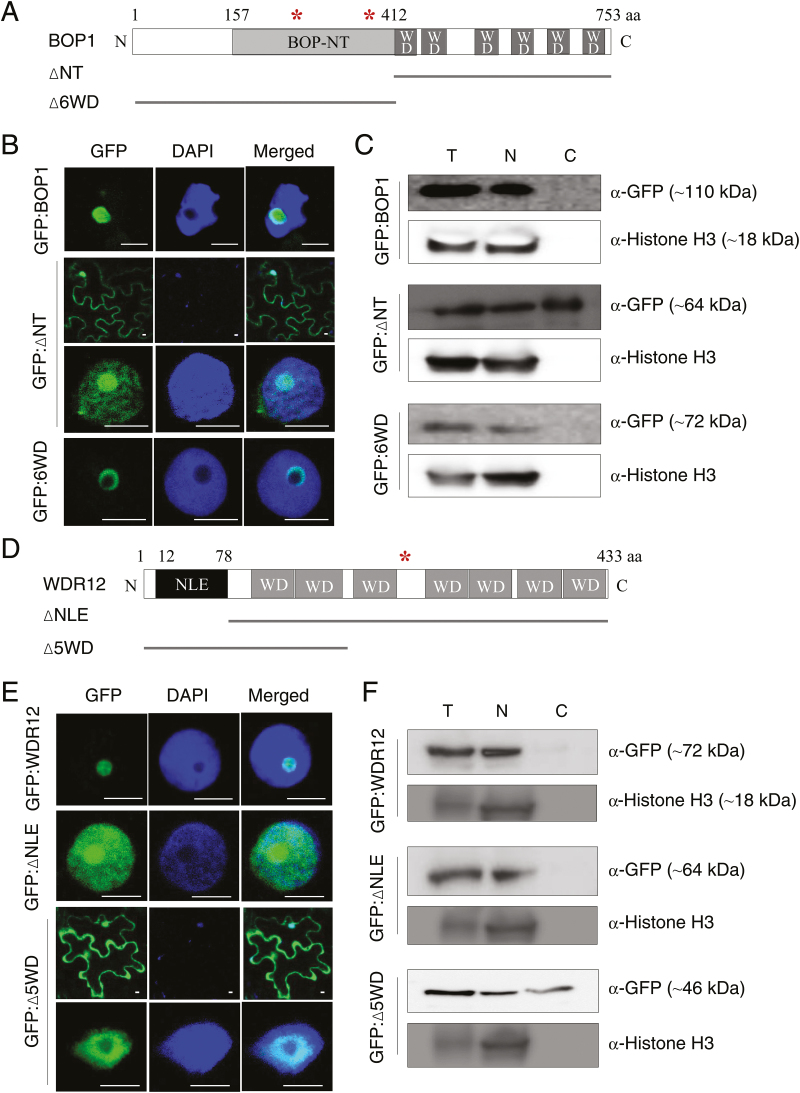
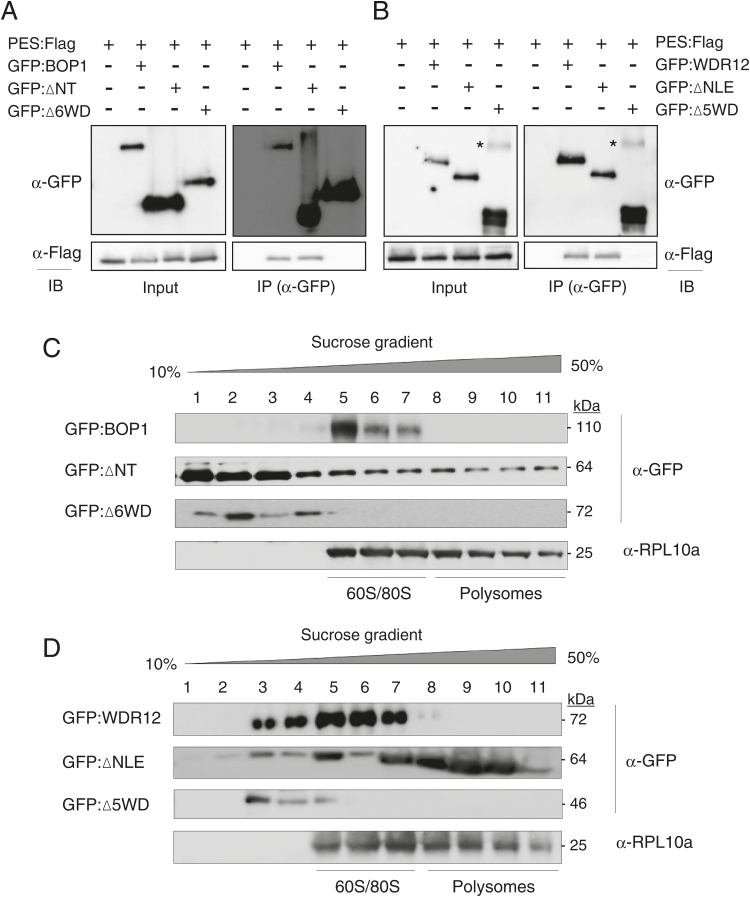
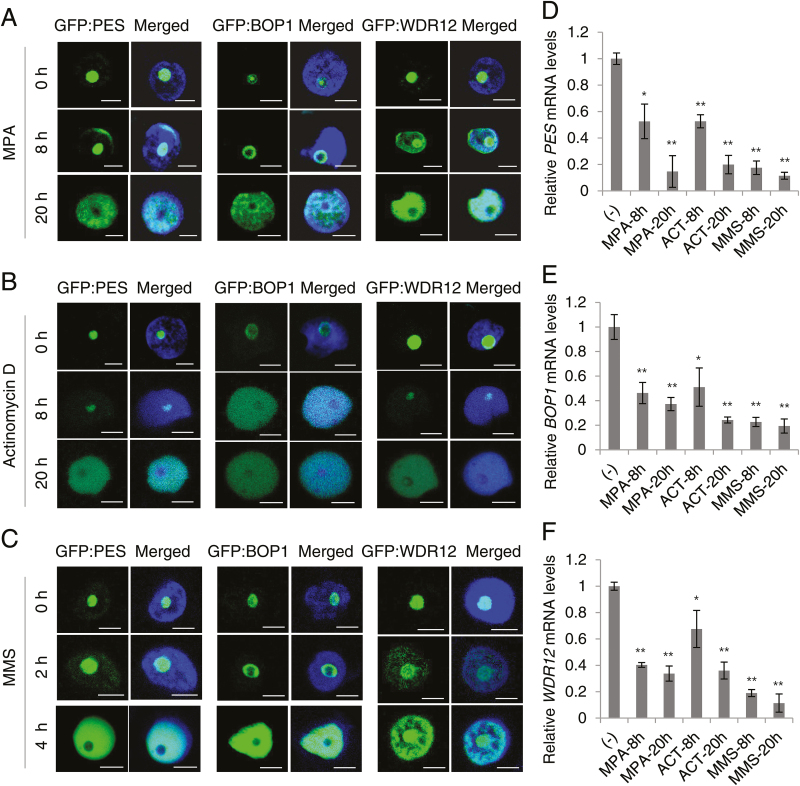
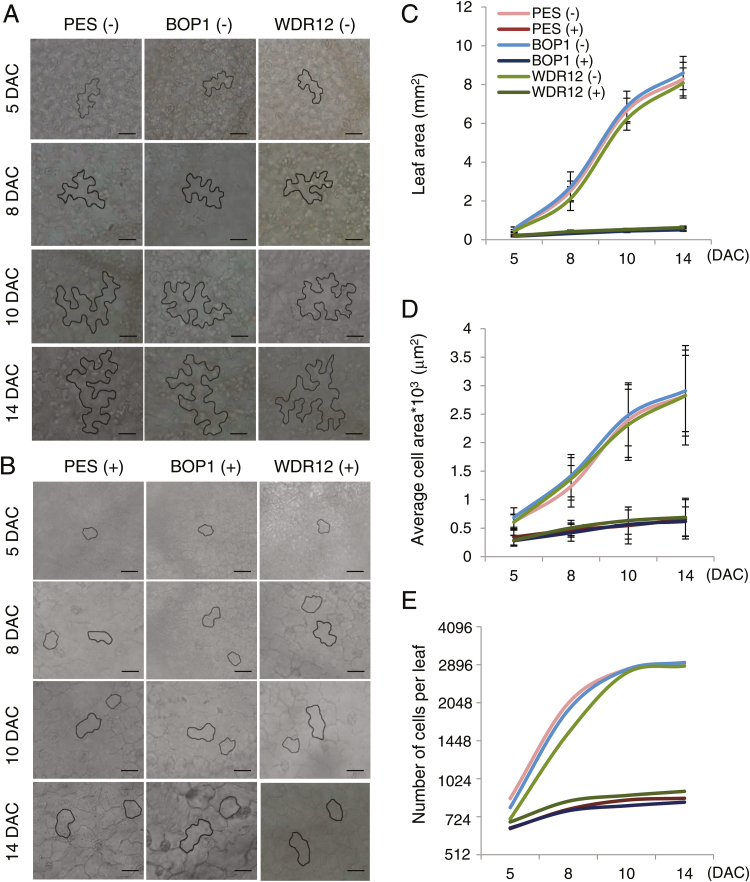
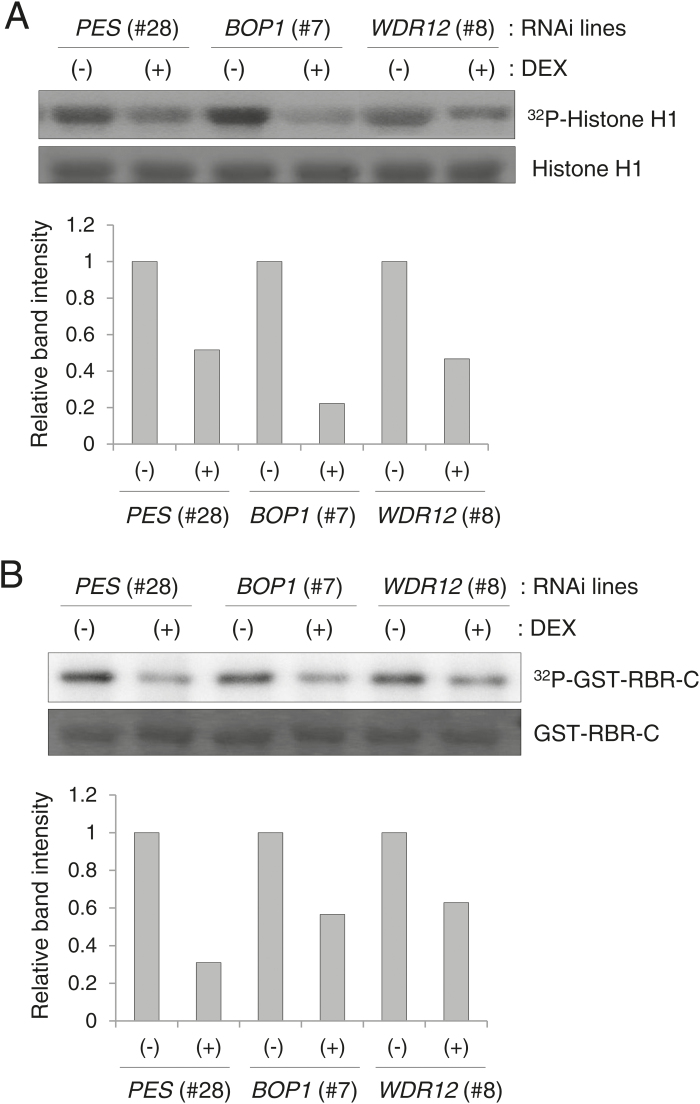
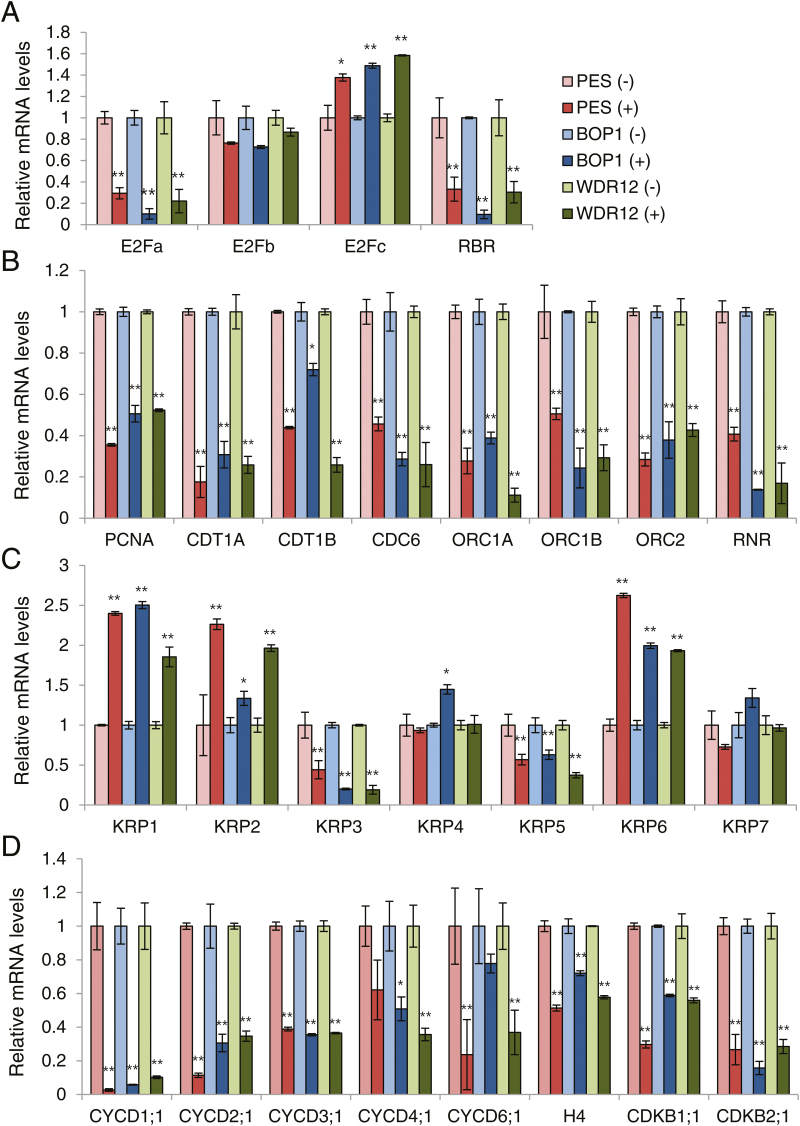
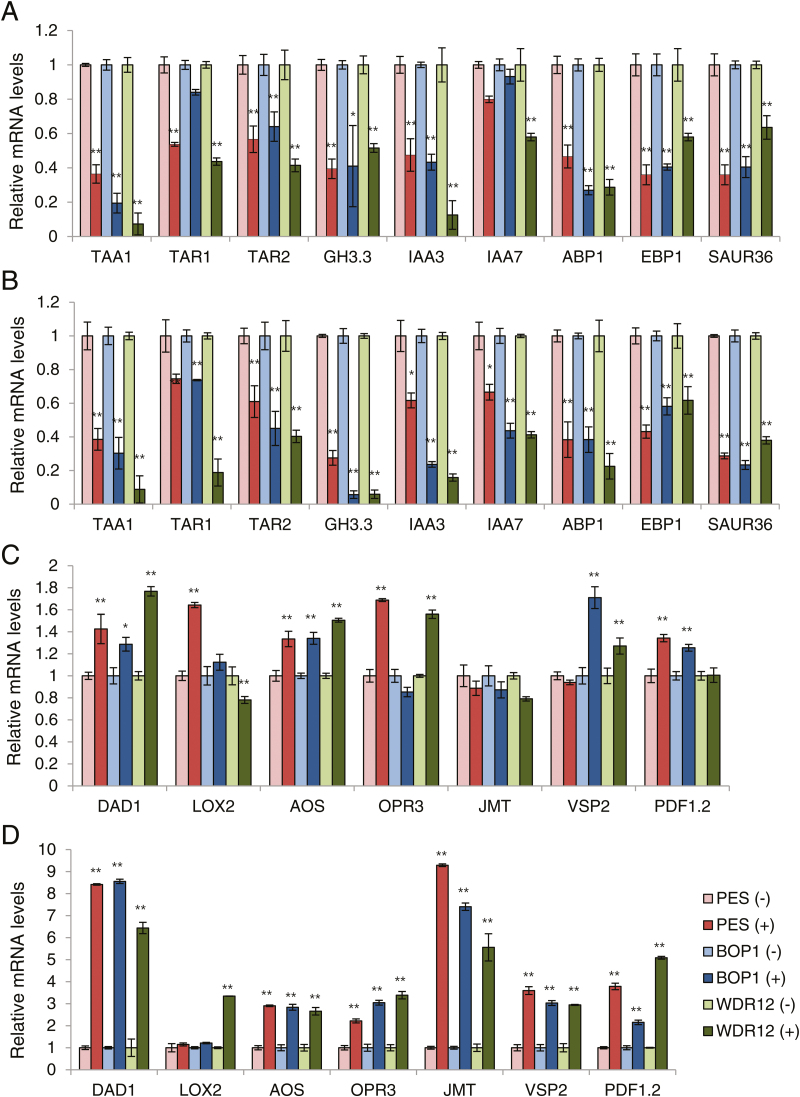
The nucleolar protein pescadillo (PES) controls biogenesis of the 60S ribosomal subunit through functional interactions with Block of Proliferation 1 (BOP1) and WD Repeat Domain 12 (WDR12) in plants. In this study, we determined protein characteristics and in planta functions of BOP1 and WDR12, and characterized defects in plant cell growth and proliferation caused by a deficiency of PeBoW (PES-BOP1-WDR12) proteins. Dexamethasone-inducible RNAi of BOP1 and WDR12 caused developmental arrest and premature senescence in Arabidopsis, similar to the phenotype of PES RNAi. Both the N-terminal domain and WD40 repeats of BOP1 and WDR12 were critical for specific associations with 60S/80S ribosomes. In response to nucleolar stress or DNA damage, PeBoW proteins moved from the nucleolus to the nucleoplasm. Kinematic analyses of leaf growth revealed that depletion of PeBoW proteins led to dramatically suppressed cell proliferation, cell expansion, and epidermal pavement cell differentiation. A deficiency in PeBoW proteins resulted in reduced cyclin-dependent kinase Type A activity, causing reduced phosphorylation of histone H1 and retinoblastoma-related (RBR) protein. PeBoW silencing caused rapid transcriptional modulation of cell-cycle genes, including reduction of E2Fa and Cyclin D family genes, and induction of several KRP genes, accompanied by down-regulation of auxin-related genes and up-regulation of jasmonic acid-related genes. Taken together, these results suggest that the PeBoW proteins involved in ribosome biogenesis play a critical role in plant cell growth and survival, and their depletion leads to inhibition of cell-cycle progression, possibly modulated by phytohormone signaling.
Keywords: Cell cycle genes; growth defects; kinematic analysis; nucleolar stress; phytohormones; ribosome biogenesis..
© The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
References
-
- Allende ML, Amsterdam A, Becker T, Kawakami K, Gaiano N, Hopkins N. 1996. Insertional mutagenesis in zebrafish identifies two novel genes, pescadillo and dead eye, essential for embryonic development. Genes & Development 10, 3141–3155. - PubMed
-
- Anastasiou E, Lenhard M. 2007. Growing up to one’s standard. Current Opinion in Plant Biology 10, 63–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
