

The Dermis as a Delivery Site of Trypanosoma brucei for Tsetse Flies
- PMID: 27441553
- PMCID: PMC4956260
- DOI: 10.1371/journal.ppat.1005744
The Dermis as a Delivery Site of Trypanosoma brucei for Tsetse Flies
Abstract
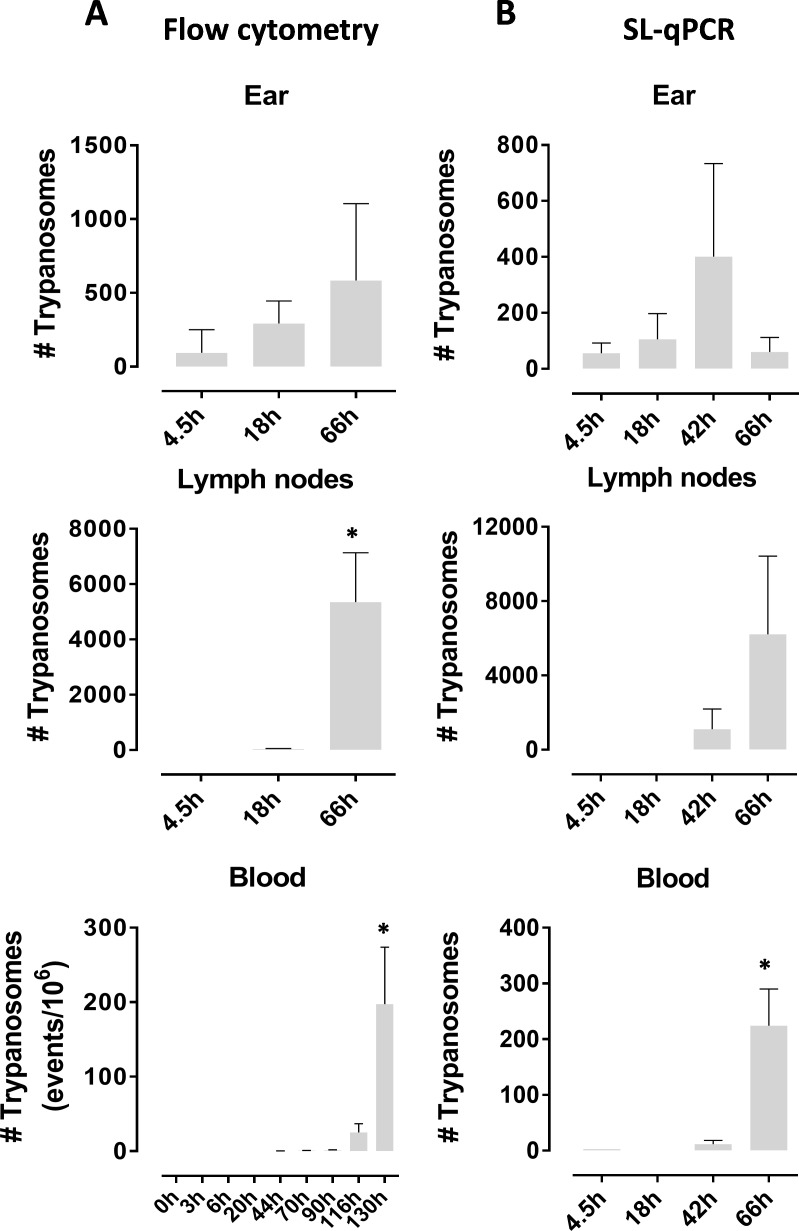
Tsetse flies are the sole vectors of Trypanosoma brucei parasites that cause sleeping sickness. Our knowledge on the early interface between the infective metacyclic forms and the mammalian host skin is currently highly limited. Glossina morsitans flies infected with fluorescently tagged T. brucei parasites were used in this study to initiate natural infections in mice. Metacyclic trypanosomes were found to be highly infectious through the intradermal route in sharp contrast with blood stream form trypanosomes. Parasite emigration from the dermal inoculation site resulted in detectable parasite levels in the draining lymph nodes within 18 hours and in the peripheral blood within 42 h. A subset of parasites remained and actively proliferated in the dermis. By initiating mixed infections with differentially labeled parasites, dermal parasites were unequivocally shown to arise from the initial inoculum and not from a re-invasion from the blood circulation. Scanning electron microscopy demonstrated intricate interactions of these skin-residing parasites with adipocytes in the connective tissue, entanglement by reticular fibers of the periadipocytic baskets and embedment between collagen bundles. Experimental transmission experiments combined with molecular parasite detection in blood fed flies provided evidence that dermal trypanosomes can be acquired from the inoculation site immediately after the initial transmission. High resolution thermographic imaging also revealed that intradermal parasite expansion induces elevated skin surface temperatures. Collectively, the dermis represents a delivery site of the highly infective metacyclic trypanosomes from which the host is systemically colonized and where a proliferative subpopulation remains that is physically constrained by intricate interactions with adipocytes and collagen fibrous structures.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
