

Face Processing Systems: From Neurons to Real-World Social Perception
- PMID: 27442071
- PMCID: PMC5345271
- DOI: 10.1146/annurev-neuro-070815-013934
Face Processing Systems: From Neurons to Real-World Social Perception
Abstract
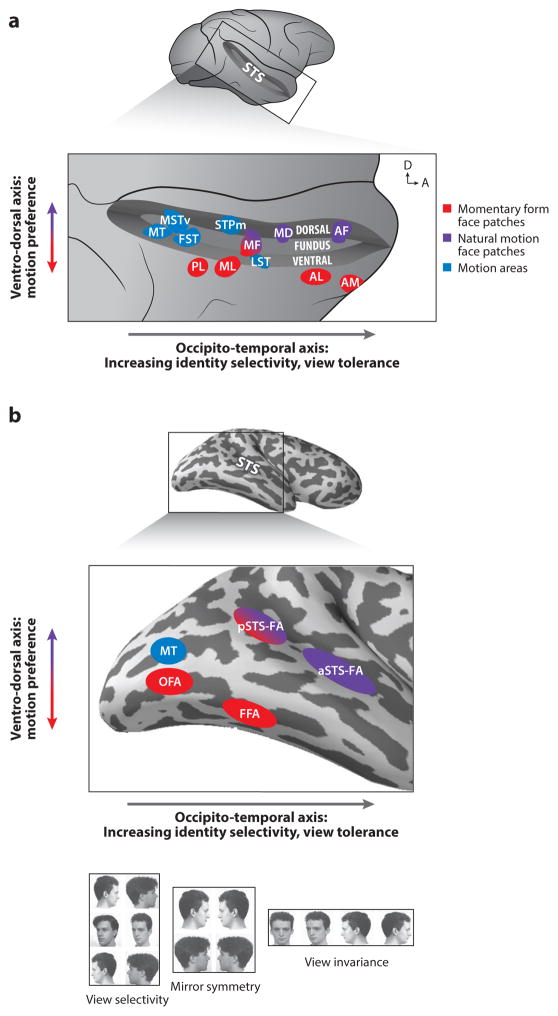
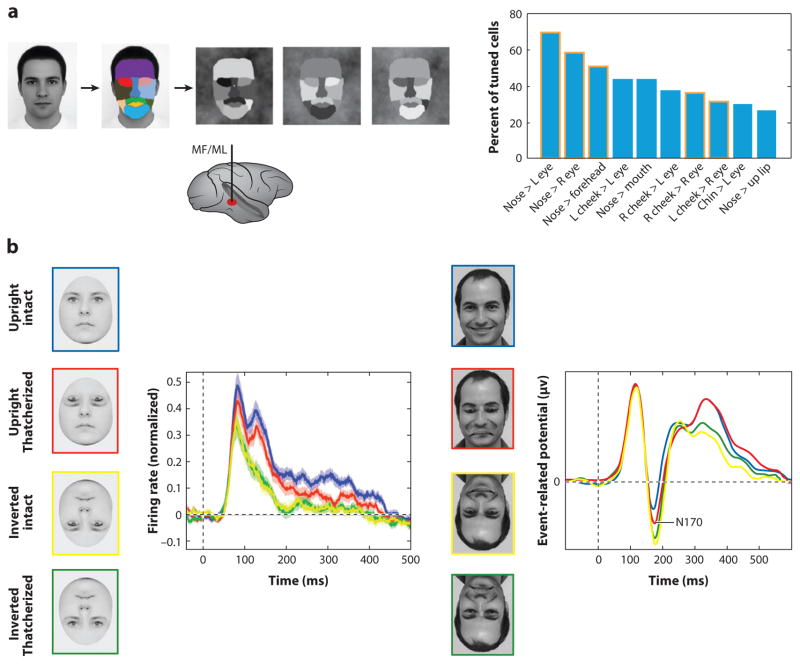
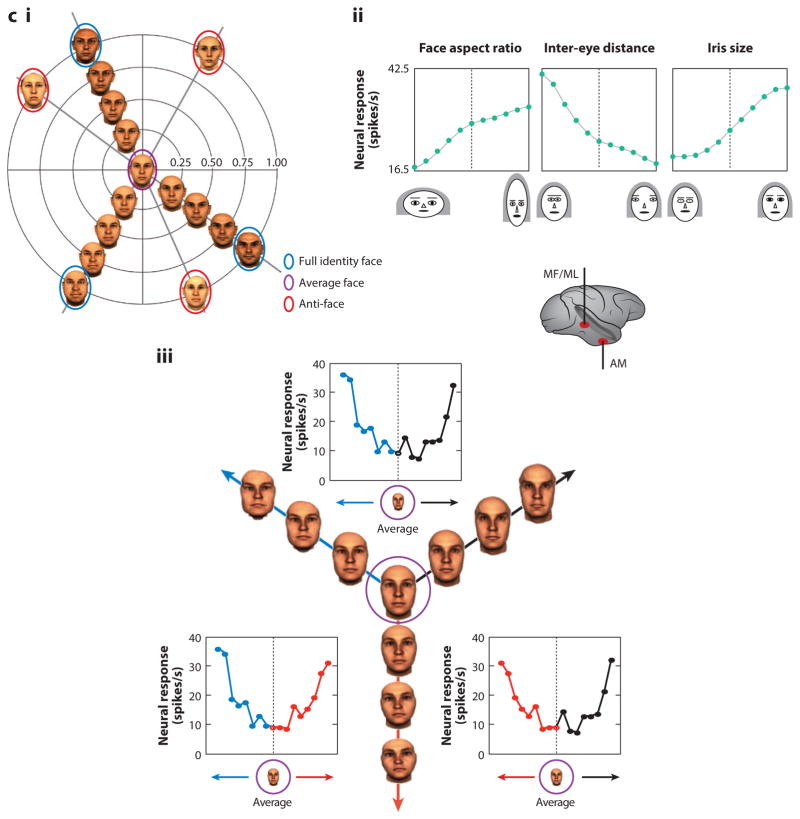
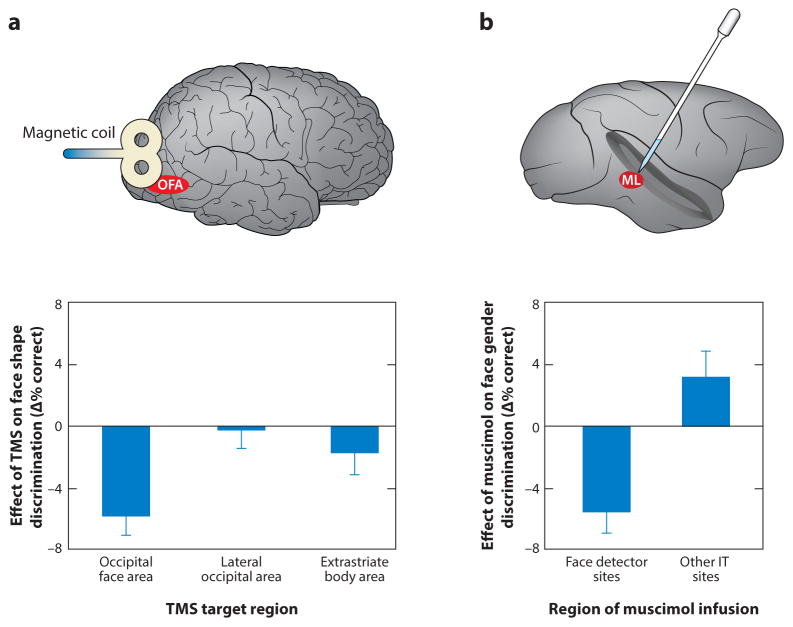
Primate face processing depends on a distributed network of interlinked face-selective areas composed of face-selective neurons. In both humans and macaques, the network is divided into a ventral stream and a dorsal stream, and the functional similarities of the areas in humans and macaques indicate they are homologous. Neural correlates for face detection, holistic processing, face space, and other key properties of human face processing have been identified at the single neuron level, and studies providing causal evidence have established firmly that face-selective brain areas are central to face processing. These mechanisms give rise to our highly accurate familiar face recognition but also to our error-prone performance with unfamiliar faces. This limitation of the face system has important implications for consequential situations such as eyewitness identification and policing.
Keywords: face recognition; functional brain organization; neural mechanisms of behavior; social brain function.
Figures
References
-
- Afraz SR, Kiani R, Esteky H. Microstimulation of inferotemporal cortex influences face categorization. Nature. 2006;442:692–5. - PubMed
-
- Andersen SM, Carlson CA, Carlson M, Gronlund SD. Individual Differences Predict Eyewitness Identification Performance. Personality and Individual Differences. 2014;60:36–40.
-
- Axelrod V, Yovel G. The challenge of localizing the anterior temporal face area: a possible solution. NeuroImage. 2013;81:371–80. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
