

A critical role for the self-assembly of Amyloid-β1-42 in neurodegeneration
- PMID: 27443509
- PMCID: PMC4957119
- DOI: 10.1038/srep30182
A critical role for the self-assembly of Amyloid-β1-42 in neurodegeneration
Abstract
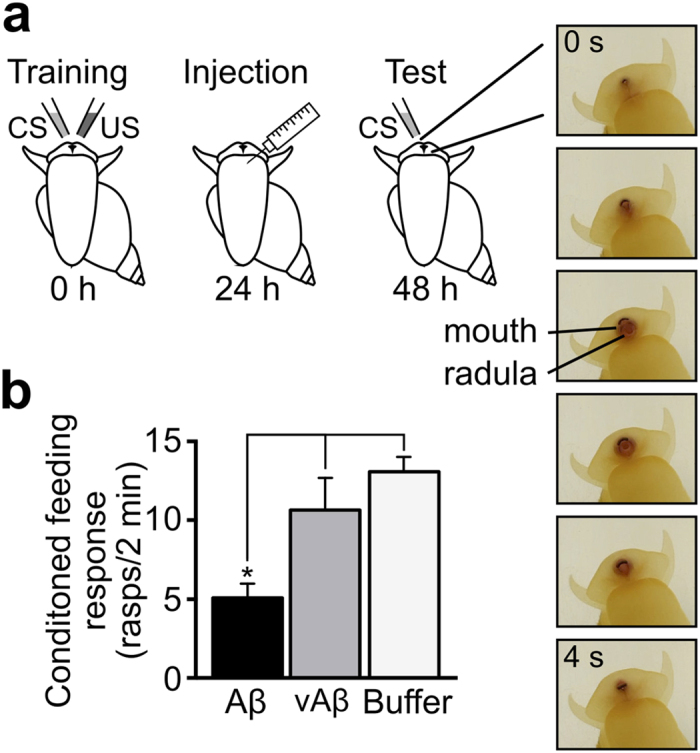
Amyloid β1-42 (Aβ1-42) plays a central role in Alzheimer's disease. The link between structure, assembly and neuronal toxicity of this peptide is of major current interest but still poorly defined. Here, we explored this relationship by rationally designing a variant form of Aβ1-42 (vAβ1-42) differing in only two amino acids. Unlike Aβ1-42, we found that the variant does not self-assemble, nor is it toxic to neuronal cells. Moreover, while Aβ1-42 oligomers impact on synaptic function, vAβ1-42 does not. In a living animal model system we demonstrate that only Aβ1-42 leads to memory deficits. Our findings underline a key role for peptide sequence in the ability to assemble and form toxic structures. Furthermore, our non-toxic variant satisfies an unmet demand for a closely related control peptide for Aβ1-42 cellular studies of disease pathology, offering a new opportunity to decipher the mechanisms that accompany Aβ1-42-induced toxicity leading to neurodegeneration.
Conflict of interest statement
The variant peptide, a kit including the variant peptide, and their uses, are covered by an International (PCT) patent application PCT/GB2015/052242 filed by the University of Sussex.
Figures





References
-
- Selkoe D. J. Toward a comprehensive theory for Alzheimer’s disease. Hypothesis: Alzheimer’s disease is caused by the cerebral accumulation and cytotoxicity of amyloid beta-protein. Annals of the New York Academy of Sciences 924, 17–25 (2000). - PubMed
-
- Hardy J. The amyloid hypothesis for Alzheimer’s disease: a critical reappraisal. J Neurochem 110, 1129–1134 (2009). - PubMed
-
- Williams T. L., Day I. J. & Serpell L. C. The effect of Alzheimer’s abeta aggregation state on the permeation of biomimetic lipid vesicles. Langmuir: the ACS journal of surfaces and colloids 26, 17260–17268 (2010). - PubMed
-
- Butterfield S. M. & Lashuel H. A. Amyloidogenic protein-membrane interactions: mechanistic insight from model systems. Angewandte Chemie 49, 5628–5654 (2010). - PubMed
-
- Shankar G. M. et al.. Natural oligomers of the Alzheimer amyloid-beta protein induce reversible synapse loss by modulating an NMDA-type glutamate receptor-dependent signaling pathway. The Journal of neuroscience: the official journal of the Society for Neuroscience 27, 2866–2875 (2007). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
