

EspR-dependent ESAT-6 Protein Secretion of Mycobacterium tuberculosis Requires the Presence of Virulence Regulator PhoP
- PMID: 27445330
- PMCID: PMC5009273
- DOI: 10.1074/jbc.M116.746289
EspR-dependent ESAT-6 Protein Secretion of Mycobacterium tuberculosis Requires the Presence of Virulence Regulator PhoP
Abstract
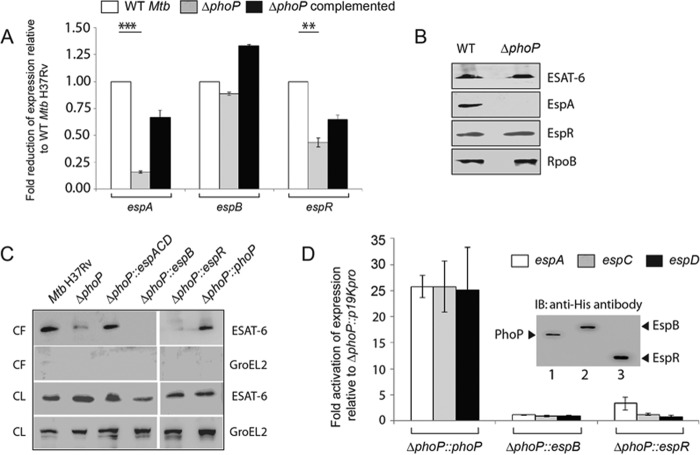
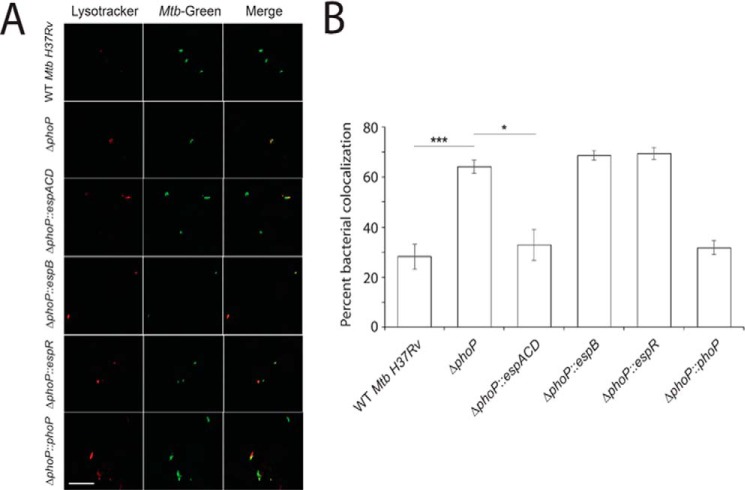
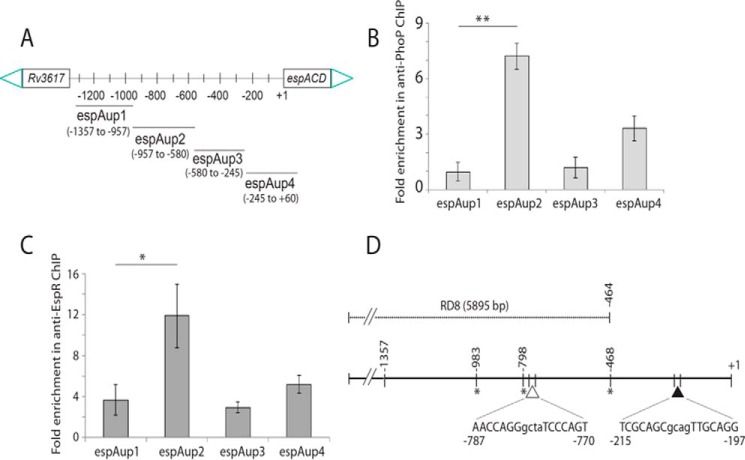
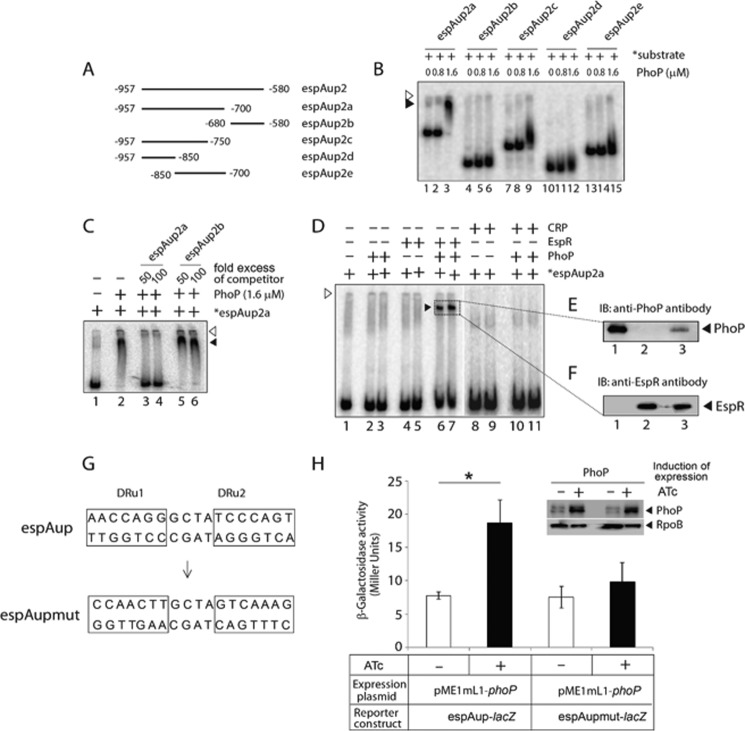
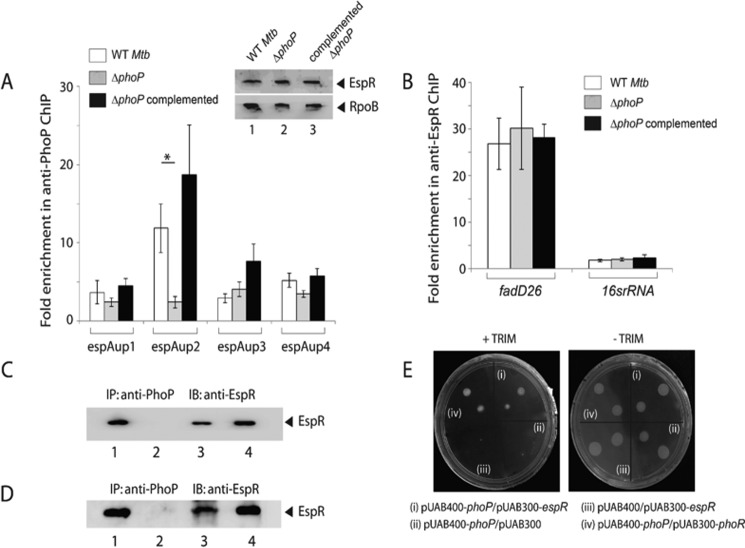
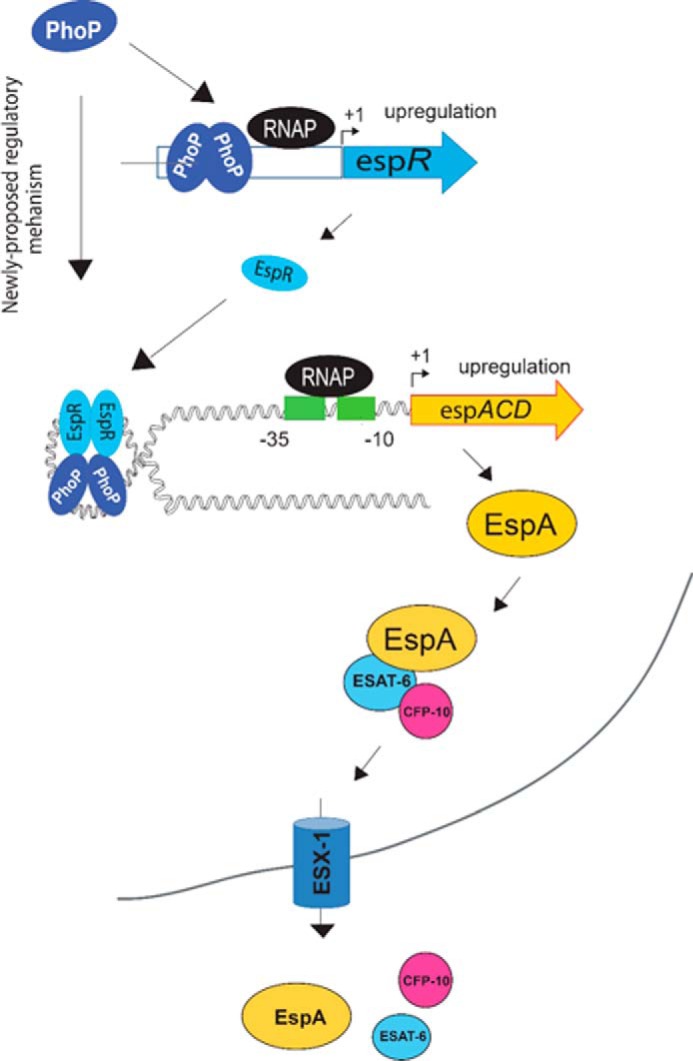
Attenuation of Mycobacterium bovis BCG strain is related to the loss of the RD1-encoded ESX-1 secretion system. The ESX-1 system secretes virulence factor ESAT-6 that plays a critical role in modulation of the host immune system, which is essential for establishment of a productive infection. Previous studies suggest that among the reasons for attenuation of Mycobacterium tuberculosis H37Ra is a mutation in the phoP gene that interferes with the ESX-1 secretion system and inhibits secretion of ESAT-6. Here, we identify a totally different and distinct regulatory mechanism involving PhoP and transcription regulator EspR on transcriptional control of the espACD operon, which is required for ESX-1-dependent ESAT-6 secretion. Although both of these regulators are capable of influencing espACD expression, we show that activation of espACD requires direct recruitment of both PhoP and EspR at the espACD promoter. The most fundamental insights are derived from the inhibition of EspR binding at the espACD regulatory region of the phoP mutant strain because of PhoP-EspR protein-protein interactions. Based on these results, a model is proposed suggesting how PhoP and EspR protein-protein interactions contribute to activation of espACD expression and, in turn, control ESAT-6 secretion, an essential pathogenic determinant of M. tuberculosis Together, these results have significant implications on the mechanism of virulence regulation of M. tuberculosis.
Keywords: DNA-protein interaction; Mycobacterium tuberculosis; gene regulation; microbial pathogenesis; protein secretion.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Bitter W., Houben E. N., Bottai D., Brodin P., Brown E. J., Cox J. S., Derbyshire K., Fortune S. M., Gao L. Y., Liu J., Gey van Pittius N. C., Pym A. S., Rubin E. J., Sherman D. R., Cole S. T., and Brosch R. (2009) Systematic genetic nomenclature for type VII secretion systems. PLoS Pathog. 5, e1000507. - PMC - PubMed
-
- Hsu T., Hingley-Wilson S. M., Chen B., Chen M., Dai A. Z., Morin P. M., Marks C. B., Padiyar J., Goulding C., Gingery M., Eisenberg D., Russell R. G., Derrick S. C., Collins F. M., Morris S. L., et al. (2003) The primary mechanism of attenuation of bacillus Calmette-Guerin is a loss of secreted lytic function required for invasion of lung interstitial tissue. Proc. Natl. Acad. Sci. U.S.A. 100, 12420–12425 - PMC - PubMed
-
- Simeone R., Bottai D., and Brosch R. (2009) ESX/type VII secretion systems and their role in host-pathogen interaction. Curr. Opin. Microbiol. 12, 4–10 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
