

Induced regulatory T cells in allograft tolerance via transient mixed chimerism
- PMID: 27446989
- PMCID: PMC4951099
- DOI: 10.1172/jci.insight.86419
Induced regulatory T cells in allograft tolerance via transient mixed chimerism
Abstract
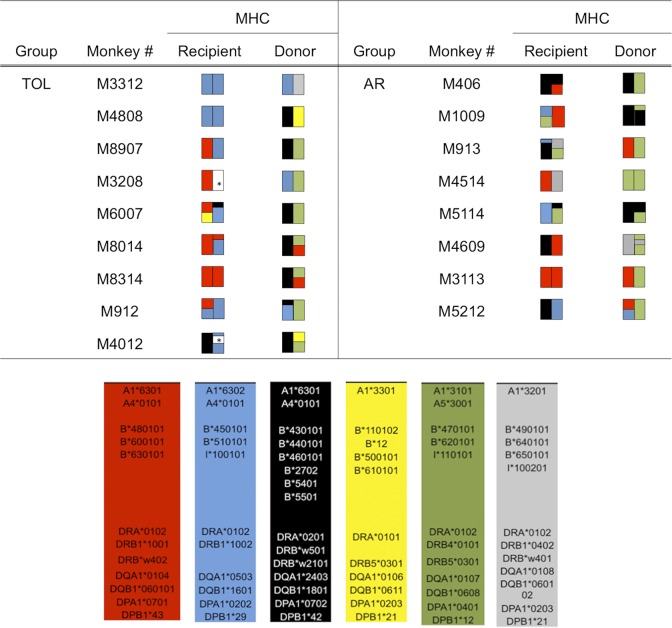
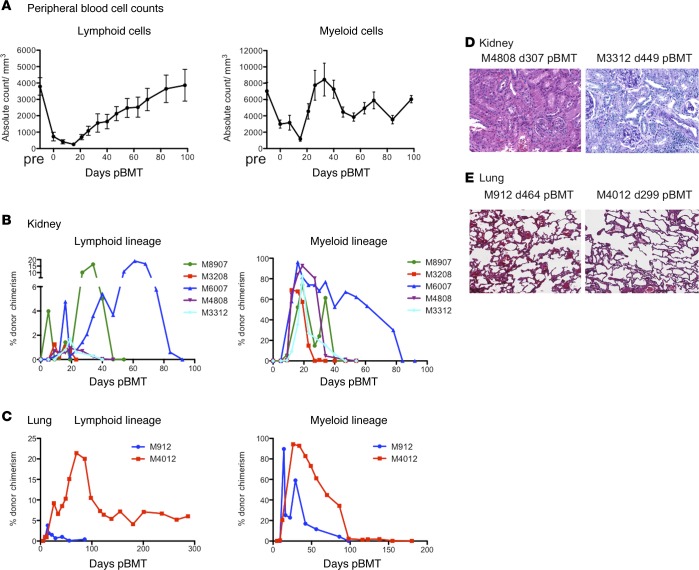
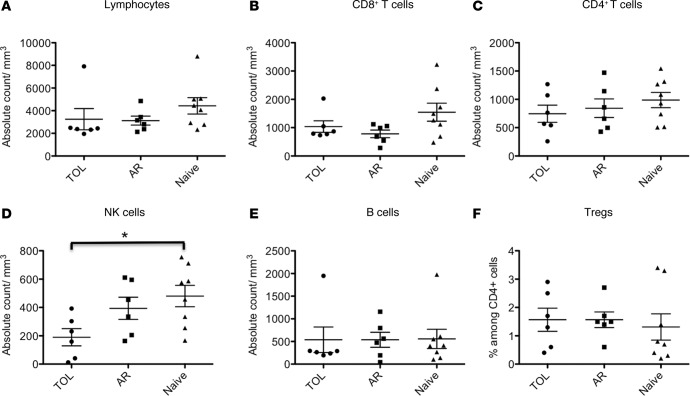
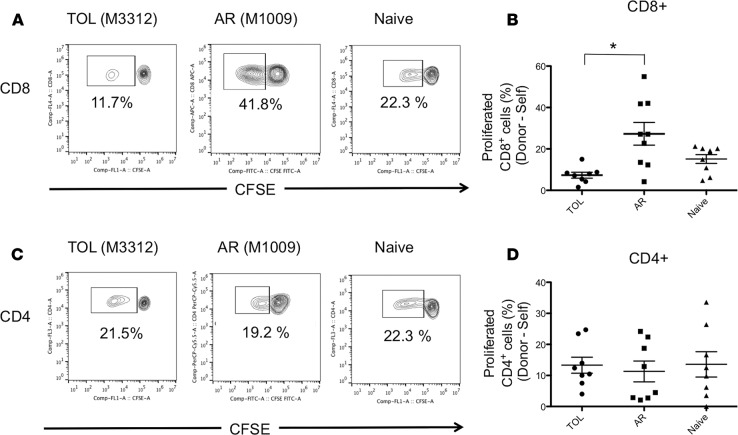
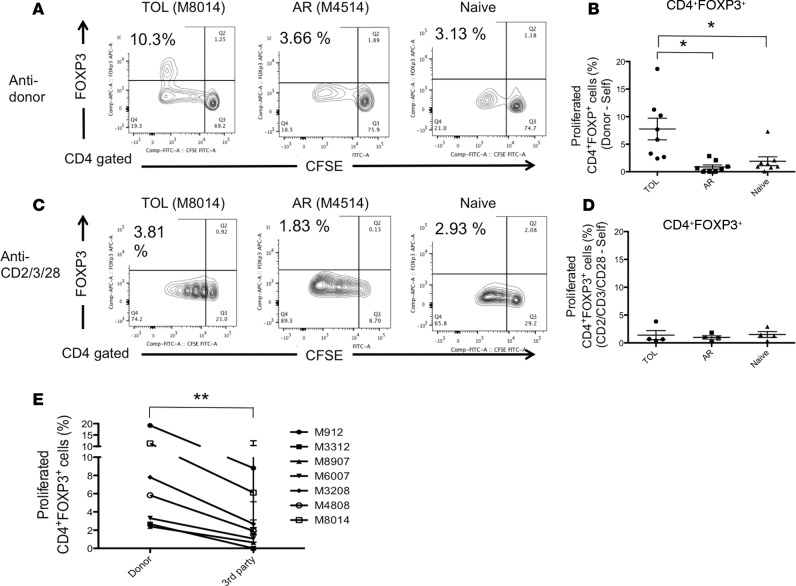
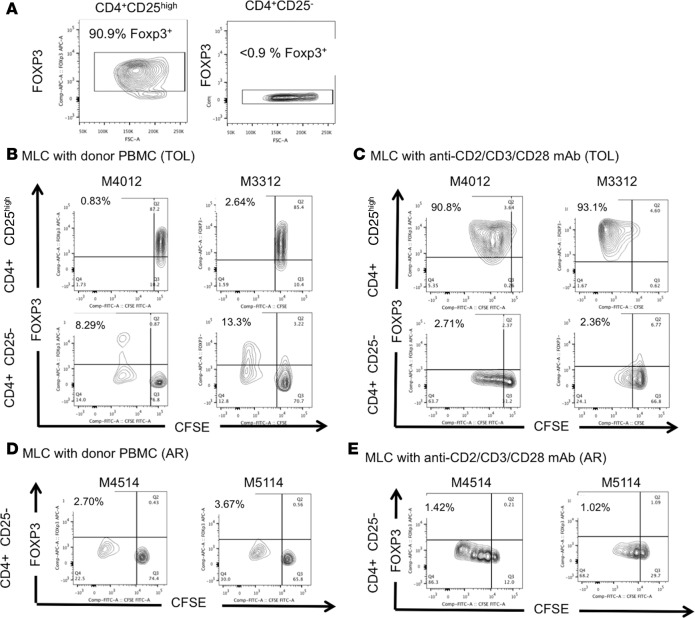
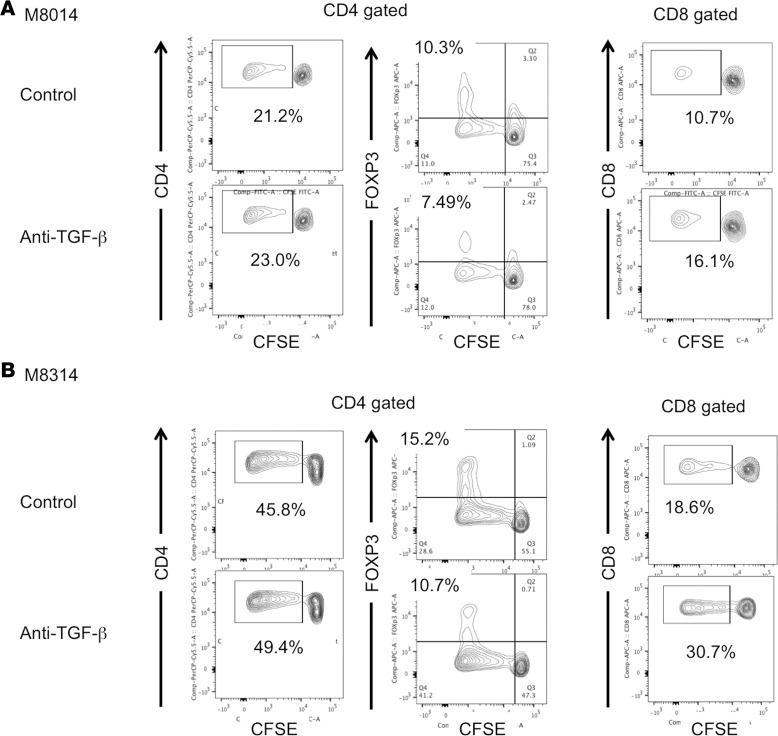
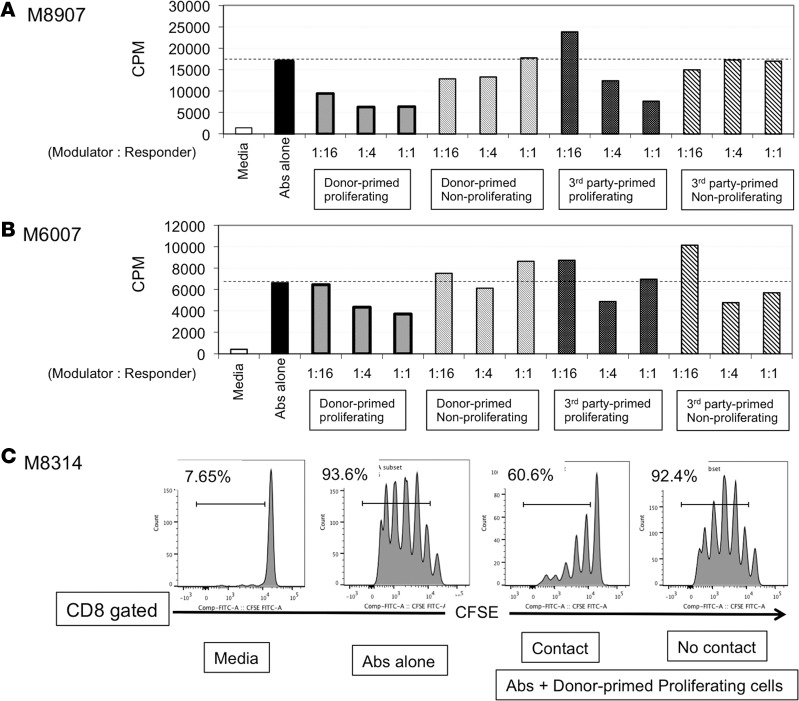
Successful induction of allograft tolerance has been achieved in nonhuman primates (NHPs) and humans via induction of transient hematopoietic chimerism. Since allograft tolerance was achieved in these recipients without durable chimerism, peripheral mechanisms are postulated to play a major role. Here, we report our studies of T cell immunity in NHP recipients that achieved long-term tolerance versus those that rejected the allograft (AR). All kidney, heart, and lung transplant recipients underwent simultaneous or delayed donor bone marrow transplantation (DBMT) following conditioning with a nonmyeloablative regimen. After DBMT, mixed lymphocyte culture with CFSE consistently revealed donor-specific loss of CD8+ T cell responses in tolerant (TOL) recipients, while marked CD4+ T cell proliferation in response to donor antigens was found to persist. Interestingly, a significant proportion of the proliferated CD4+ cells were FOXP3+ in TOL recipients, but not in AR or naive NHPs. In TOL recipients, CD4+FOXP3+ cell proliferation against donor antigens was greater than that observed against third-party antigens. Finally, the expanded Tregs appeared to be induced Tregs (iTregs) that were converted from non-Tregs. These data provide support for the hypothesis that specific induction of iTregs by donor antigens is key to long-term allograft tolerance induced by transient mixed chimerism.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
