

Target-dependent biogenesis of cognate microRNAs in human cells
- PMID: 27448149
- PMCID: PMC4961841
- DOI: 10.1038/ncomms12200
Target-dependent biogenesis of cognate microRNAs in human cells
Abstract
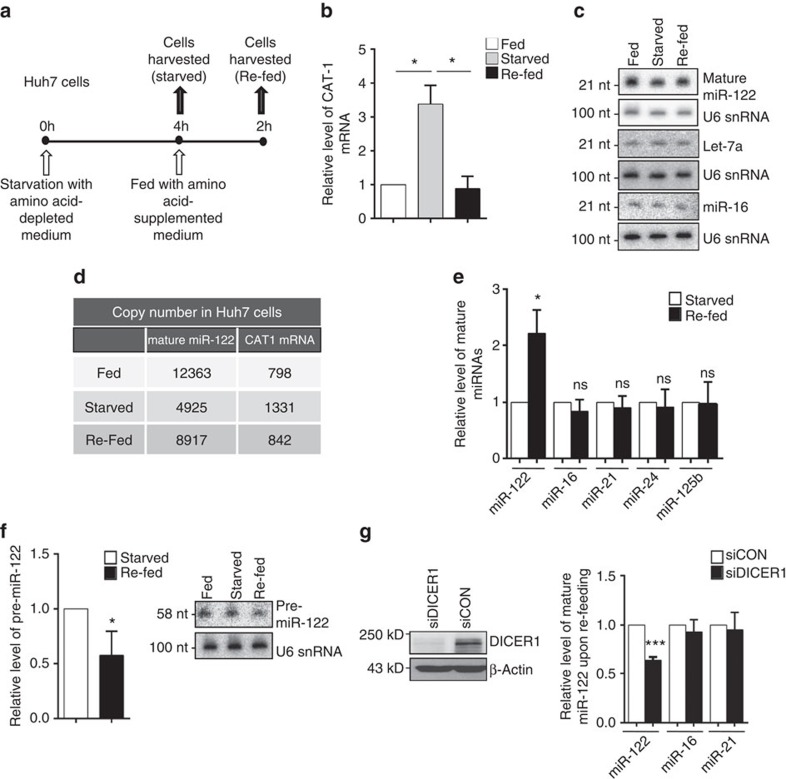
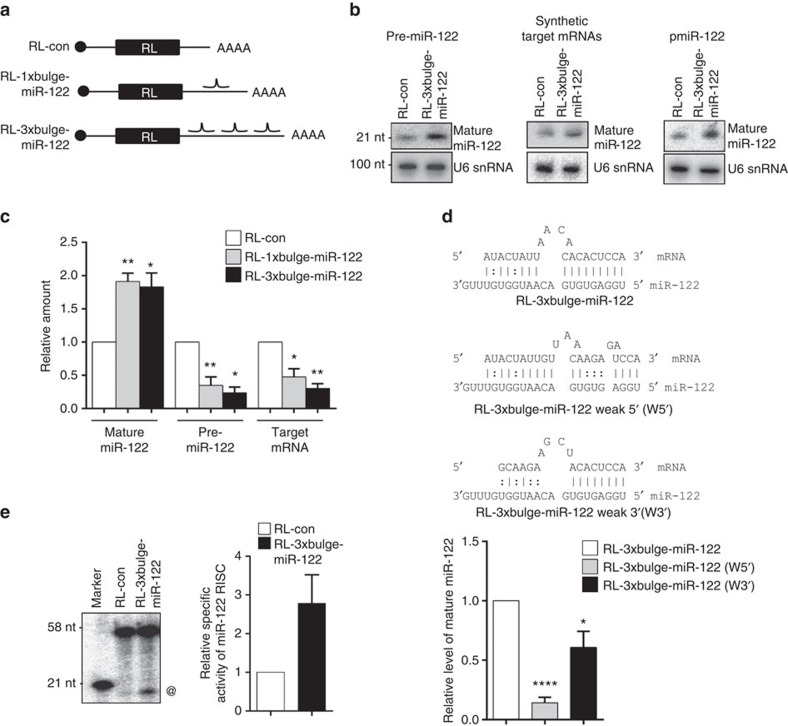
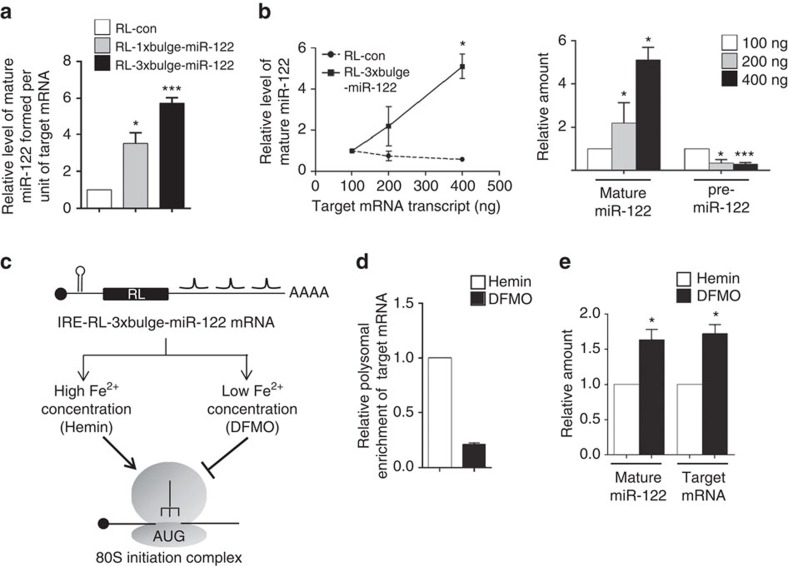
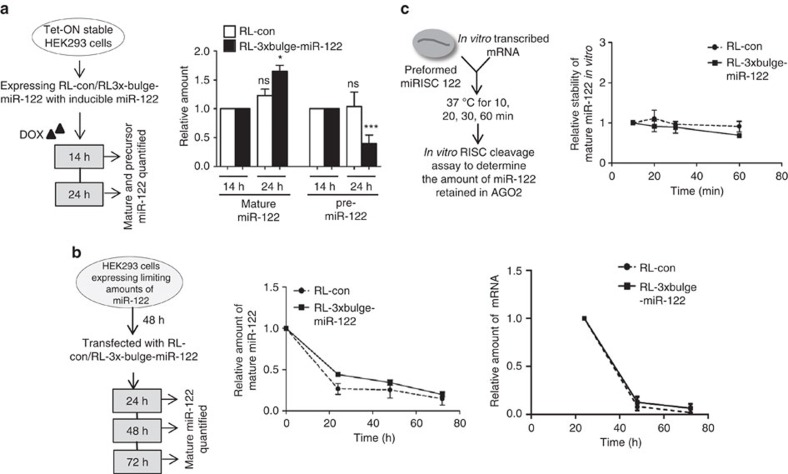
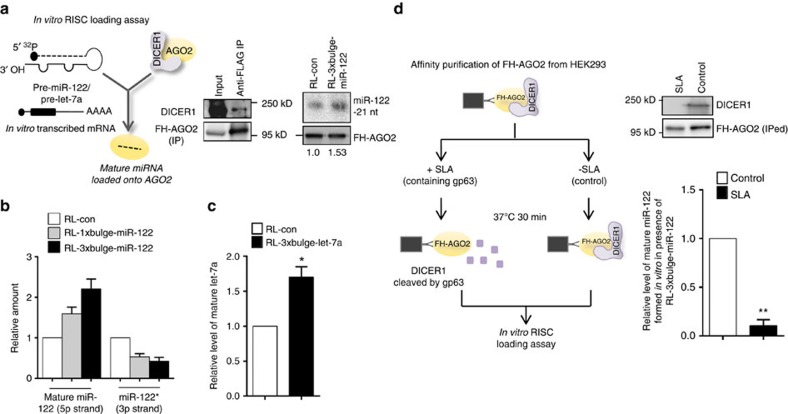
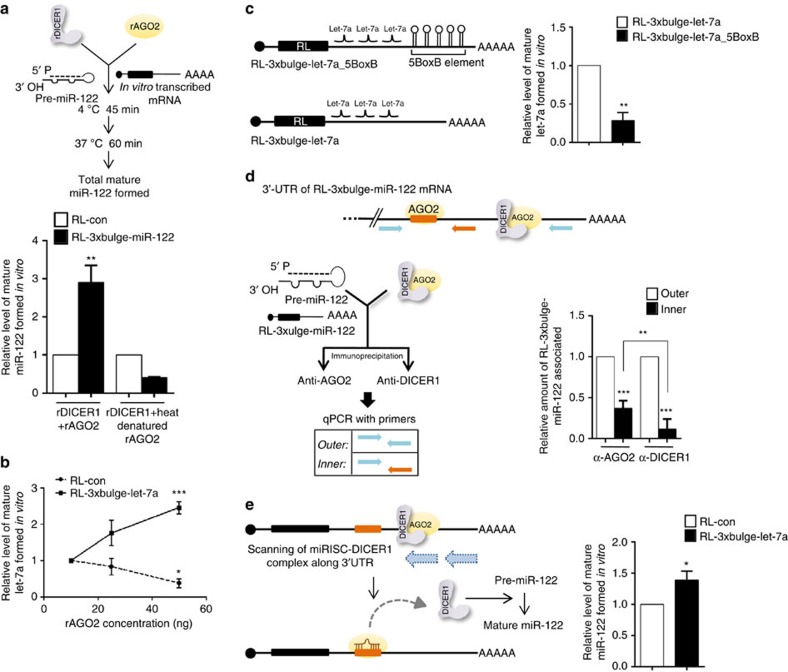
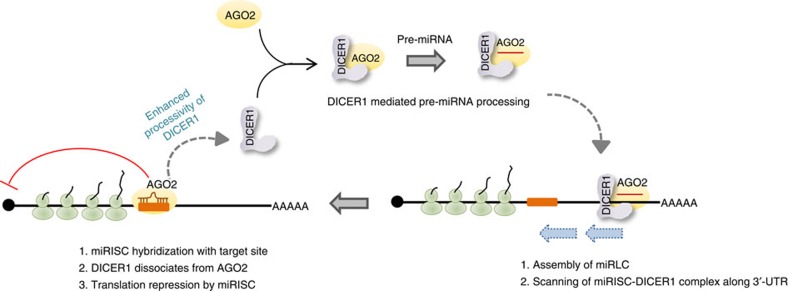
Extensive research has established how miRNAs regulate target mRNAs by translation repression and/or endonucleolytic degradation in metazoans. However, information related to the effect of target mRNA on biogenesis and stability of corresponding miRNAs in animals is limited. Here we report regulated biogenesis of cognate miRNAs by their target mRNAs. Enhanced pre-miRNA processing by AGO-associated DICER1 contributes to this increased miRNP formation. The processed miRNAs are loaded onto AGO2 to form functionally competent miRISCs both in vivo and also in a cell-free in vitro system. Thus, we identify an additional layer of posttranscriptional regulation that helps the cell to maintain requisite levels of mature forms of respective miRNAs by modulating their processing in a target-dependent manner, a process happening for miR-122 during stress reversal in human hepatic cells.
Conflict of interest statement
The authors declare no conflict of interests.
Figures
References
-
- Ameres S. L., Martinez J. & Schroeder R. Molecular basis for target RNA recognition and cleavage by human RISC. Cell 130, 101–112 (2007). - PubMed
-
- Sasaki T., Shiohama A., Minoshima S. & Shimizu N. Identification of eight members of the Argonaute family in the human genome. Genomics 82, 323–330 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
