

Prolonged Particulate Hexavalent Chromium Exposure Suppresses Homologous Recombination Repair in Human Lung Cells
- PMID: 27449664
- PMCID: PMC5601504
- DOI: 10.1093/toxsci/kfw103
Prolonged Particulate Hexavalent Chromium Exposure Suppresses Homologous Recombination Repair in Human Lung Cells
Abstract
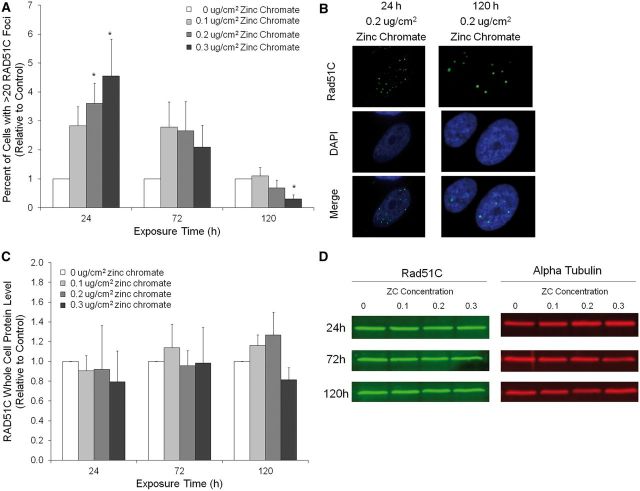
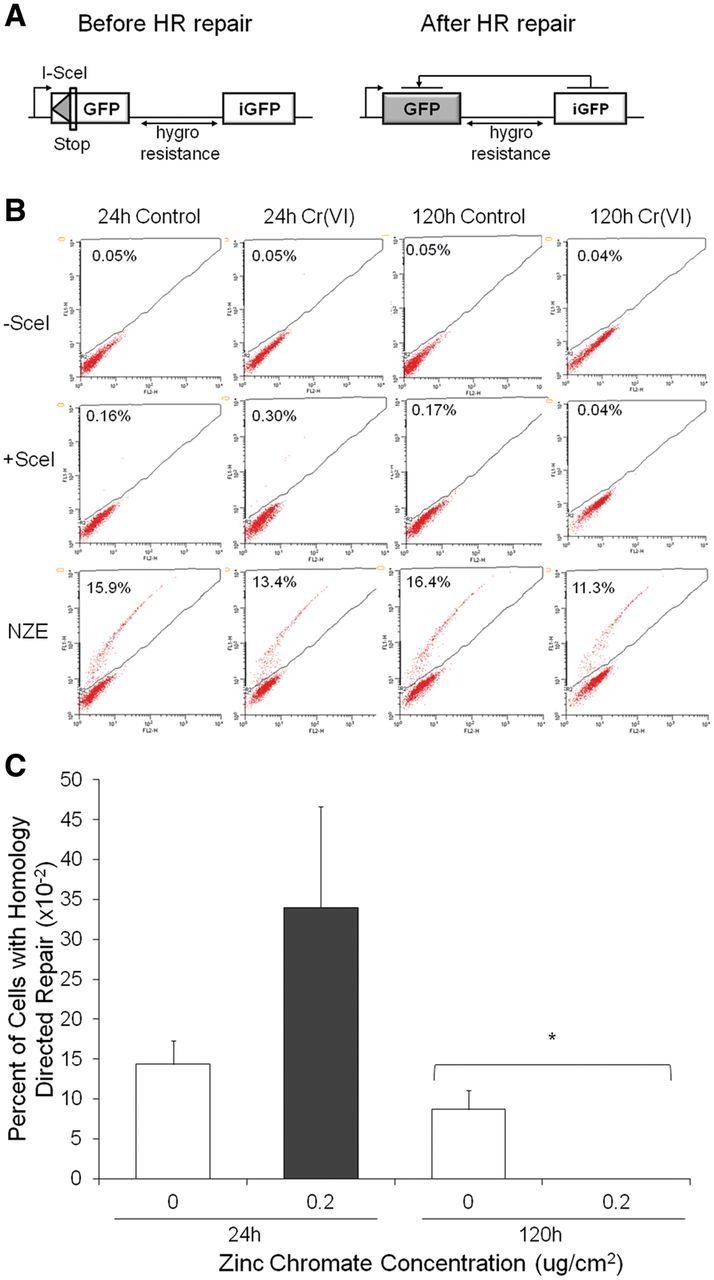
Genomic instability is one of the primary models of carcinogenesis and a feature of almost all cancers. Homologous recombination (HR) repair protects against genomic instability by maintaining high genomic fidelity during the repair of DNA double strand breaks. The defining step of HR repair is the formation of the Rad51 nucleofilament, which facilitates the search for a homologous sequence and invasion of the template DNA strand. Particulate hexavalent chromium (Cr(VI)), a human lung carcinogen, induces DNA double strand breaks and chromosome instability. Since the loss of HR repair increases Cr(VI)-induced chromosome instability, we investigated the effect of extended Cr(VI) exposure on HR repair. We show acute (24 h) Cr(VI) exposure induces a normal HR repair response. In contrast, prolonged (120 h) exposure to particulate Cr(VI) inhibited HR repair and Rad51 nucleofilament formation. Prolonged Cr(VI) exposure had a profound effect on Rad51, evidenced by reduced protein levels and Rad51 mislocalization to the cytoplasm. The response of proteins involved in Rad51 nuclear import and nucleofilament formation displayed varying responses to prolonged Cr(VI) exposure. BRCA2 formed nuclear foci after prolonged Cr(VI) exposure, while Rad51C foci formation was suppressed. These results suggest that particulate Cr(VI), a major chemical carcinogen, inhibits HR repair by targeting Rad51, causing DNA double strand breaks to be repaired by a low fidelity, Rad51-independent repair pathway. These results further enhance our understanding of the underlying mechanism of Cr(VI)-induced chromosome instability and thus, carcinogenesis.
Keywords: Rad51 nucleofilament; chromosome instability; homologous recombination repair; particulate chromium (VI).
© The Author 2016. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures
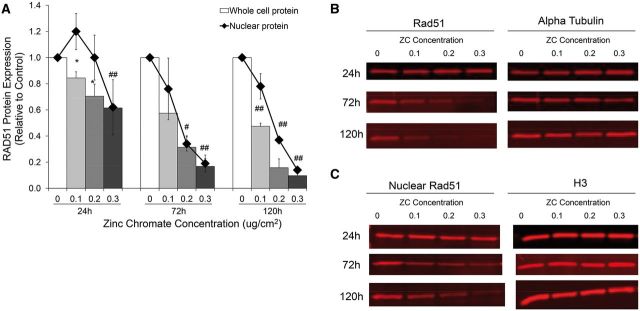
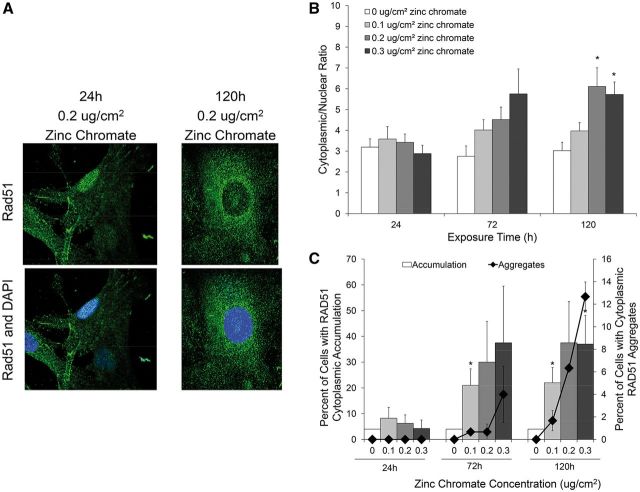
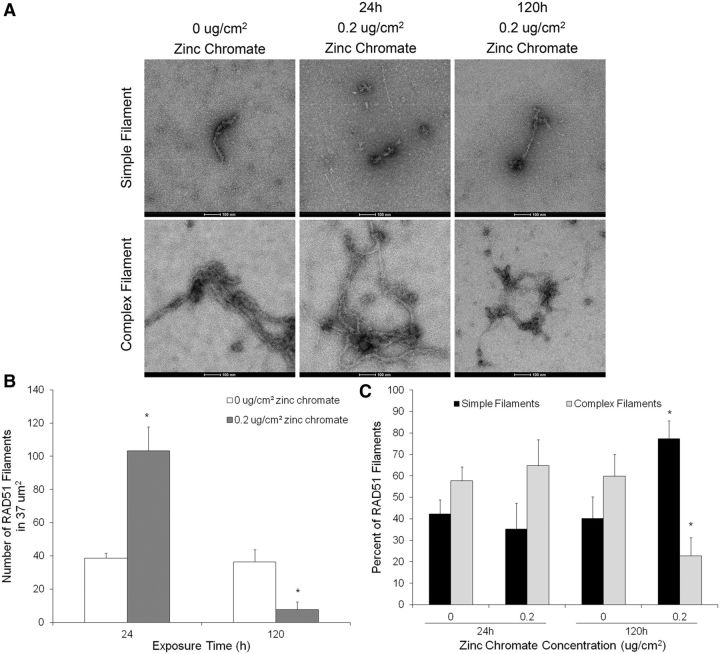
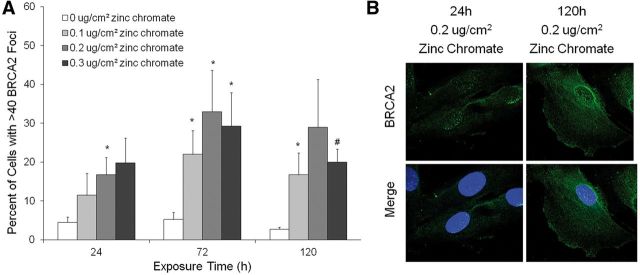
References
-
- Baumann P., Benson F. E., West S. C. (1996). Human Rad51 protein promotes ATP-dependent homologous pairing and strand transfer reactions in vitro. Cell 87, 757–766. - PubMed
-
- Bryant H. E., Ying S., Helleday T. (2006). Homologous recombination is involved in repair of chromium-induced DNA damage in mammalian cells. Mutat. Res. 599, 116–123. - PubMed
-
- Candelli A., Holthausen J. T., Depken M., Brouwer I., Franker M.A.M., Marchetti M., Heller I., Bernard S., Garcin E. B., Modesti M., et al. (2014). Visualization and quantification of nascent Rad41 filament formation at single-moner resolution. Proc. Natl. Acad. Sci U S A 111, 15090–15095. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
