

Orexin A attenuates palmitic acid-induced hypothalamic cell death
- PMID: 27449757
- PMCID: PMC5399885
- DOI: 10.1016/j.mcn.2016.07.003
Orexin A attenuates palmitic acid-induced hypothalamic cell death
Abstract
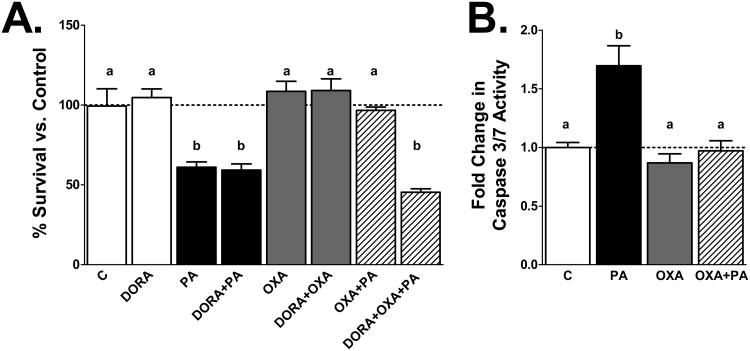
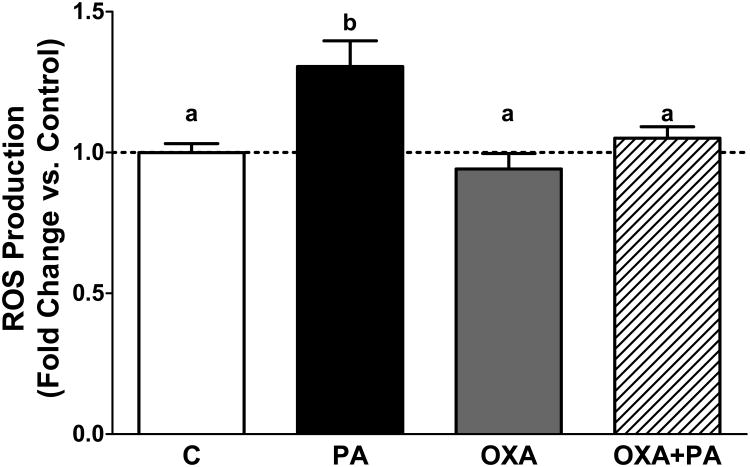
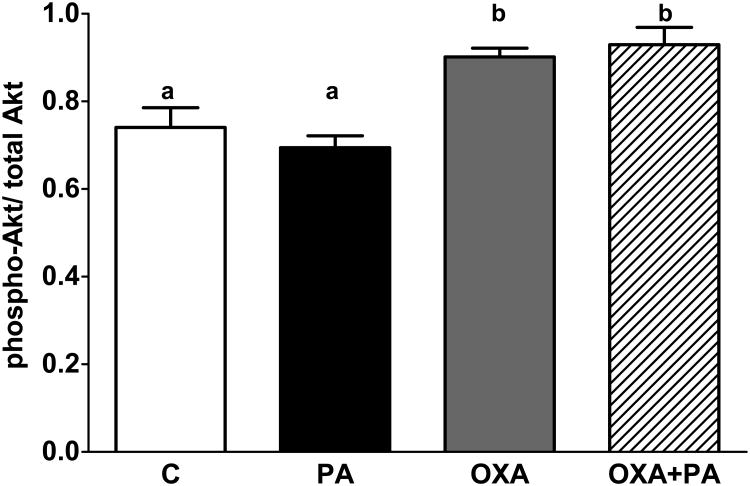
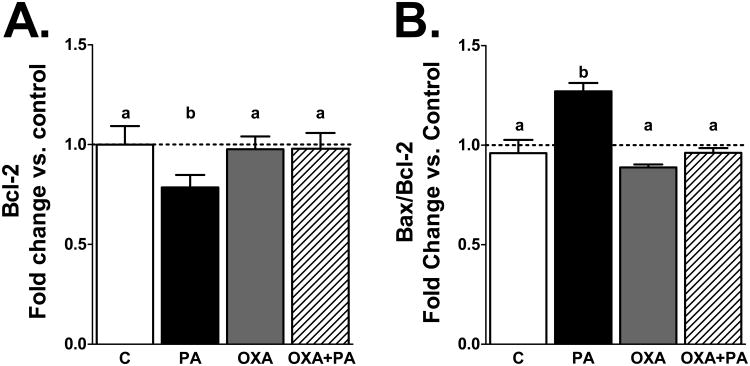
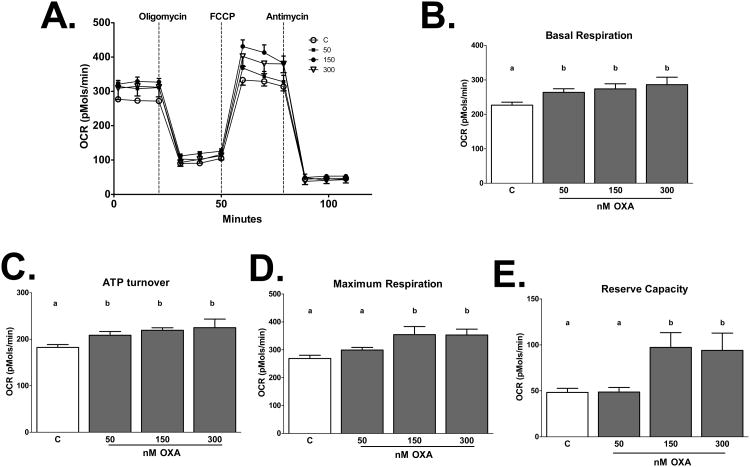
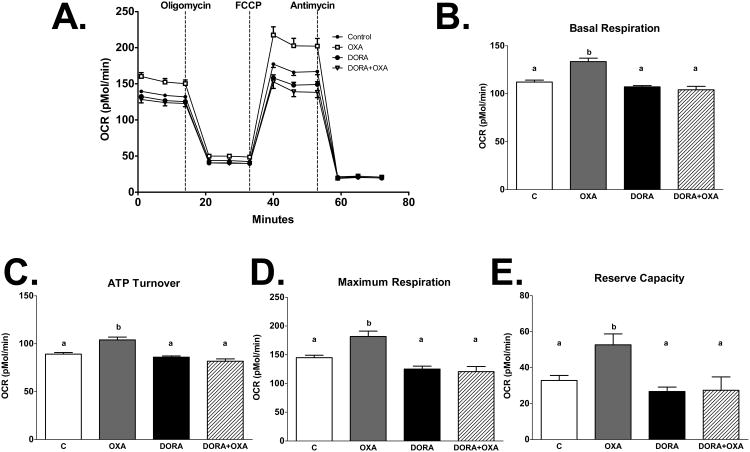
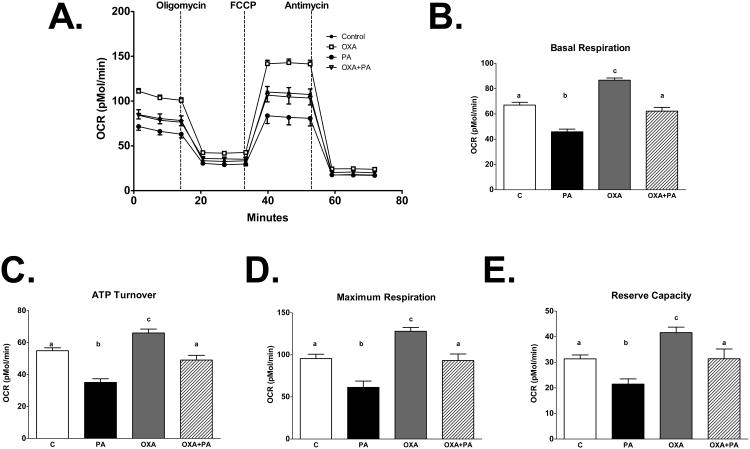
Palmitic acid (PA), an abundant dietary saturated fatty acid, contributes to obesity and hypothalamic dysregulation in part through increase in oxidative stress, insulin resistance, and neuroinflammation. Increased production of reactive oxygen species (ROS) as a result of PA exposure contributes to the onset of neuronal apoptosis. Additionally, high fat diets lead to changes in hypothalamic gene expression profiles including suppression of the anti-apoptotic protein B cell lymphoma 2 (Bcl-2) and upregulation of the pro-apoptotic protein B cell lymphoma 2 associated X protein (Bax). Orexin A (OXA), a hypothalamic peptide important in obesity resistance, also contributes to neuroprotection. Prior studies have demonstrated that OXA attenuates oxidative stress induced cell death. We hypothesized that OXA would be neuroprotective against PA induced cell death. To test this, we treated an immortalized hypothalamic cell line (designated mHypoA-1/2) with OXA and PA. We demonstrate that OXA attenuates PA-induced hypothalamic cell death via reduced caspase-3/7 apoptosis, stabilization of Bcl-2 gene expression, and reduced Bax/Bcl-2 gene expression ratio. We also found that OXA inhibits ROS production after PA exposure. Finally, we show that PA exposure in mHypoA-1/2 cells significantly reduces basal respiration, maximum respiration, ATP production, and reserve capacity. However, OXA treatment reverses PA-induced changes in intracellular metabolism, increasing basal respiration, maximum respiration, ATP production, and reserve capacity. Collectively, these results support that OXA protects against PA-induced hypothalamic dysregulation, and may represent one mechanism through which OXA can ameliorate effects of obesogenic diet on brain health.
Keywords: Apoptosis; Hypocretin; Neurodegeneration; Neuroprotection; Palmitic acid; Reactive oxygen species.
Published by Elsevier Inc.
Figures
Similar articles
-
Role of orexin A signaling in dietary palmitic acid-activated microglial cells.Neurosci Lett. 2015 Oct 8;606:140-4. doi: 10.1016/j.neulet.2015.08.033. Epub 2015 Aug 22. Neurosci Lett. 2015. PMID: 26306651 Free PMC article.
-
Orexin A decreases lipid peroxidation and apoptosis in a novel hypothalamic cell model.Neurosci Lett. 2012 Aug 22;524(1):30-4. doi: 10.1016/j.neulet.2012.07.002. Epub 2012 Jul 11. Neurosci Lett. 2012. PMID: 22796468 Free PMC article.
-
Melatonin pre-treatment mitigates SHSY-5Y cells against oxaliplatin induced mitochondrial stress and apoptotic cell death.PLoS One. 2017 Jul 21;12(7):e0180953. doi: 10.1371/journal.pone.0180953. eCollection 2017. PLoS One. 2017. PMID: 28732061 Free PMC article.
-
Protection Strategies Against Palmitic Acid-Induced Lipotoxicity in Metabolic Syndrome and Related Diseases.Int J Mol Sci. 2025 Jan 18;26(2):788. doi: 10.3390/ijms26020788. Int J Mol Sci. 2025. PMID: 39859502 Free PMC article. Review.
-
Role of the saturated fatty acid palmitate in the interconnected hypothalamic control of energy homeostasis and biological rhythms.Am J Physiol Endocrinol Metab. 2018 Aug 1;315(2):E133-E140. doi: 10.1152/ajpendo.00433.2017. Epub 2018 Apr 6. Am J Physiol Endocrinol Metab. 2018. PMID: 29631363 Review.
Cited by
-
Functional consequences of brain exposure to saturated fatty acids: From energy metabolism and insulin resistance to neuronal damage.Endocrinol Diabetes Metab. 2023 Jan;6(1):e386. doi: 10.1002/edm2.386. Epub 2022 Nov 2. Endocrinol Diabetes Metab. 2023. PMID: 36321333 Free PMC article. Review.
-
Synchronous neuronal interactions in rat hypothalamic culture: a novel model for the study of network dynamics in metabolic disorders.Exp Brain Res. 2021 Mar;239(3):755-764. doi: 10.1007/s00221-020-05977-7. Epub 2021 Jan 3. Exp Brain Res. 2021. PMID: 33388905
-
Orexin A-Mediated Modulation of Reproductive Activities in Testis of Normal and Cryptorchid Dogs: Possible Model for Studying Relationships Between Energy Metabolism and Reproductive Control.Front Endocrinol (Lausanne). 2019 Nov 22;10:816. doi: 10.3389/fendo.2019.00816. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31824429 Free PMC article.
-
Mitochondrial-Respiration-Improving Effects of Three Different Gardeniae Fructus Preparations and Their Components.ACS Omega. 2021 Dec 7;6(50):34229-34241. doi: 10.1021/acsomega.1c03265. eCollection 2021 Dec 21. ACS Omega. 2021. PMID: 34963909 Free PMC article.
-
Endocrinopathies and Male Infertility.Life (Basel). 2021 Dec 22;12(1):10. doi: 10.3390/life12010010. Life (Basel). 2021. PMID: 35054403 Free PMC article. Review.
References
-
- Belsham DD, Fick LJ, Dalvi PS, Centeno ML, Chalmers JA, Lee PK, Wang Y, Drucker DJ, Koletar MM. Ciliary neurotrophic factor recruitment of glucagon-like peptide-1 mediates neurogenesis, allowing immortalization of adult murine hypothalamic neurons. Faseb J. 2009;23:4256–4265. doi: 10.1096/fj.09-133454. - DOI - PubMed
-
- Benoit SC, Kemp CJ, Elias CF, Abplanalp W, Herman JP, Migrenne S, Lefevre AL, Cruciani-Guglielmacci C, Magnan C, Yu F, Niswender K, Irani BG, Holland WL, Clegg DJ. Palmitic acid mediates hypothalamic insulin resistance by altering PKC-theta subcellular localization in rodents. The Journal of clinical investigation. 2009;119:2577–2589. doi: 10.1172/JCI36714. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
