

Volvox: A simple algal model for embryogenesis, morphogenesis and cellular differentiation
- PMID: 27451296
- PMCID: PMC5101179
- DOI: 10.1016/j.ydbio.2016.07.014
Volvox: A simple algal model for embryogenesis, morphogenesis and cellular differentiation
Abstract
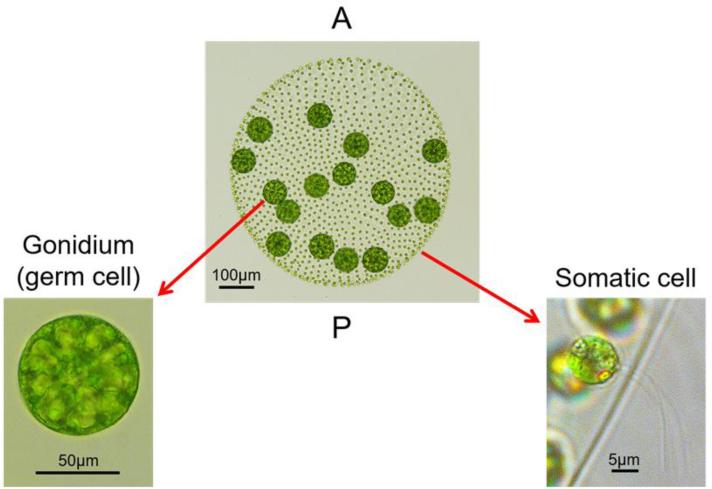
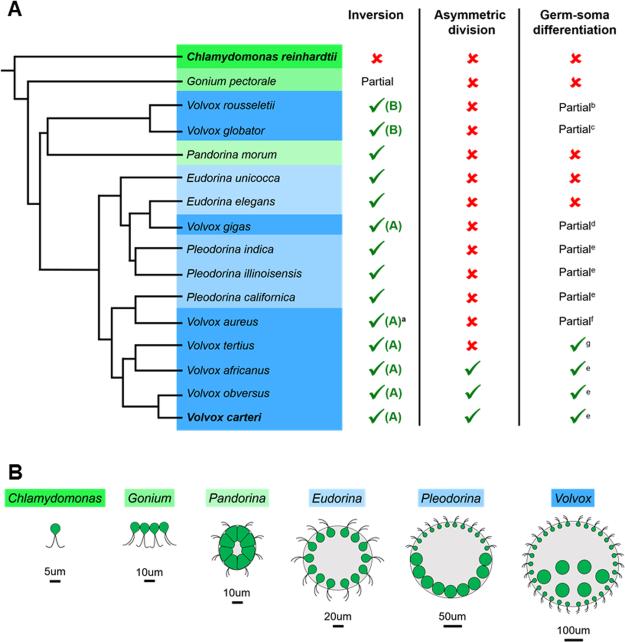
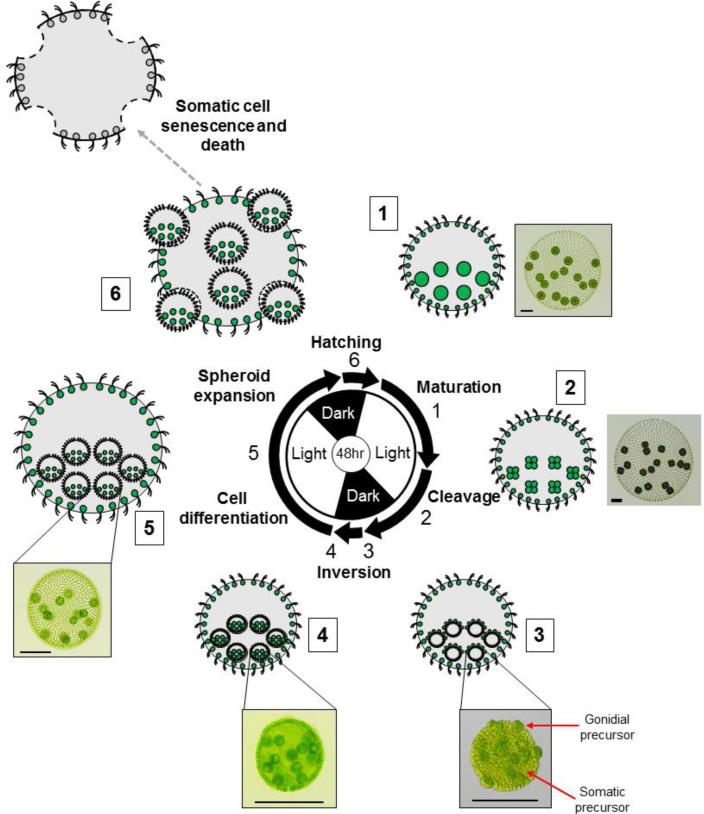
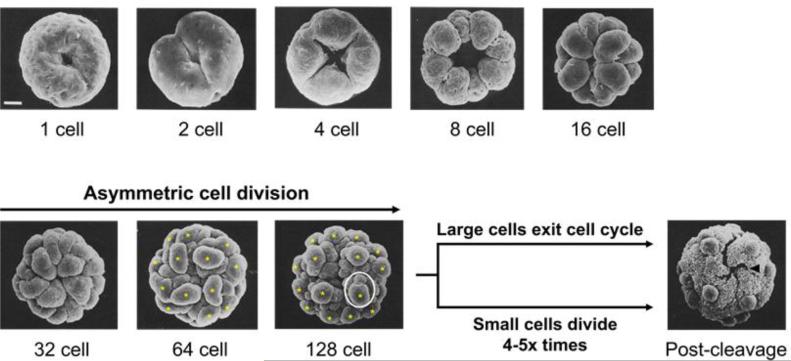
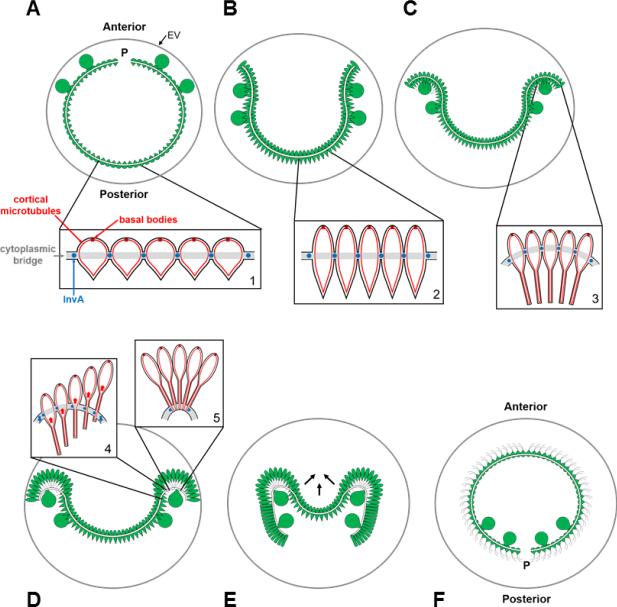
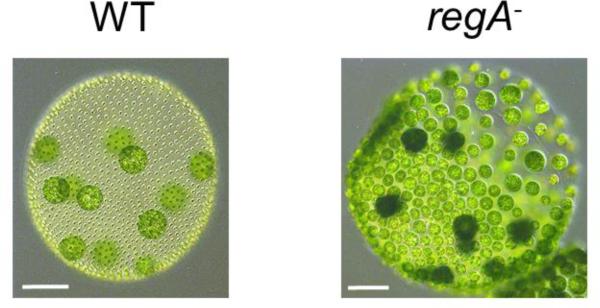
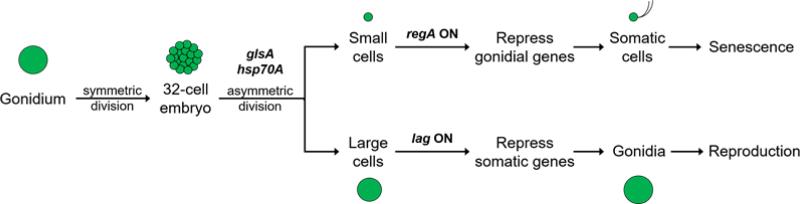
Patterning of a multicellular body plan involves a coordinated set of developmental processes that includes cell division, morphogenesis, and cellular differentiation. These processes have been most intensively studied in animals and land plants; however, deep insight can also be gained by studying development in simpler multicellular organisms. The multicellular green alga Volvox carteri (Volvox) is an excellent model for the investigation of developmental mechanisms and their evolutionary origins. Volvox has a streamlined body plan that contains only a few thousand cells and two distinct cell types: reproductive germ cells and terminally differentiated somatic cells. Patterning of the Volvox body plan is achieved through a stereotyped developmental program that includes embryonic cleavage with asymmetric cell division, morphogenesis, and cell-type differentiation. In this review we provide an overview of how these three developmental processes give rise to the adult form in Volvox and how developmental mutants have provided insights into the mechanisms behind these events. We highlight the accessibility and tractability of Volvox and its relatives that provide a unique opportunity for studying development.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Bell G. The origin and early evolution of germ cells as illustrated by the Volvocales. In: Halvorson H, Monroy A, editors. The Origin and Evolution of Sex. Wood's Hole Symposium. Liss, New York: 1985. pp. 221–256.
-
- Benfey PN. Essays on Developmental Biology, Part A. Elsevier Inc.; 2016. Defining the Path from Stem Cells to Differentiated Tissue; pp. 35–43. doi:10.1016/bs.ctdb.2015.12.002. - PubMed
-
- Bišová K, Zachleder V. Cell-cycle regulation in green algae dividing by multiple fission. J. Exp. Bot. 2014;65:2585–602. doi:10.1093/jxb/ert466. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
