

Beta Cell Hubs Dictate Pancreatic Islet Responses to Glucose
- PMID: 27452146
- PMCID: PMC5031557
- DOI: 10.1016/j.cmet.2016.06.020
Beta Cell Hubs Dictate Pancreatic Islet Responses to Glucose
Abstract
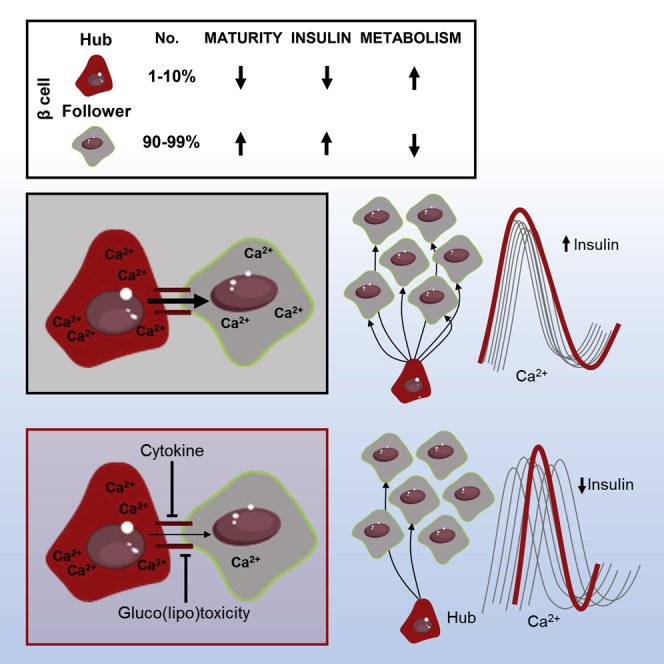
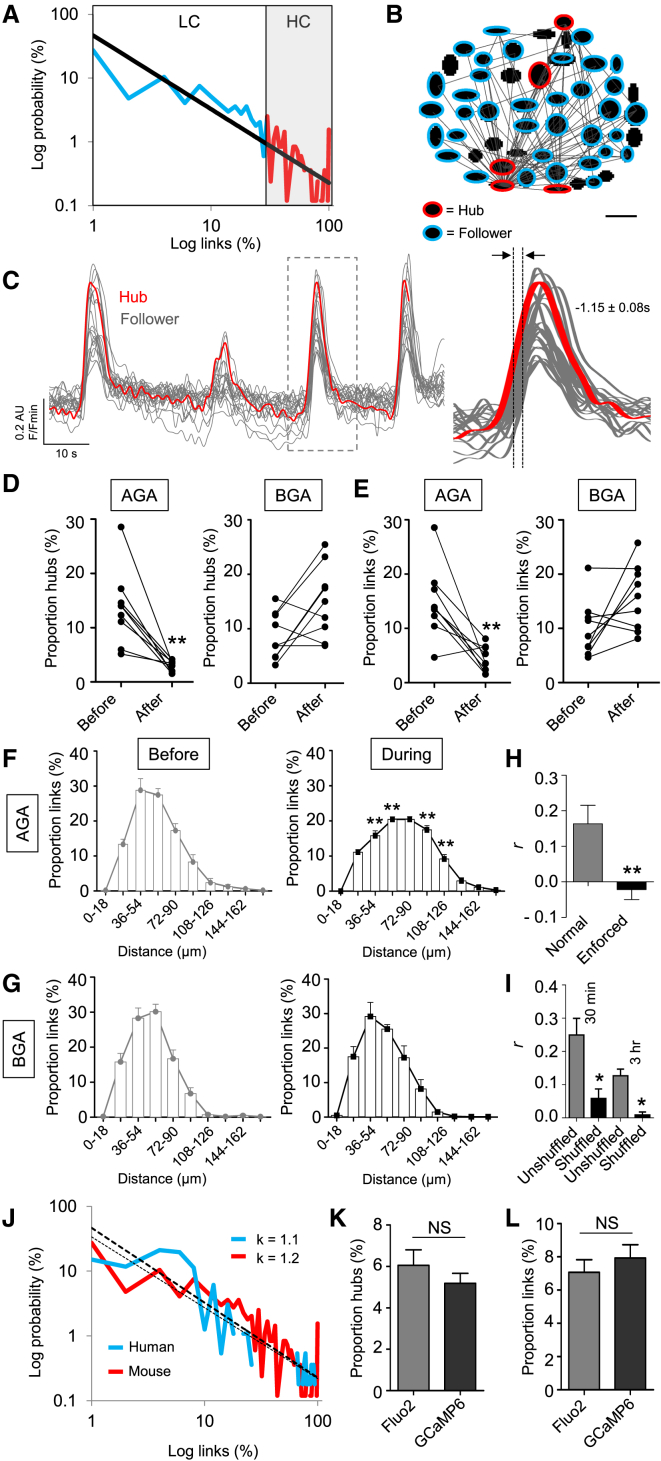
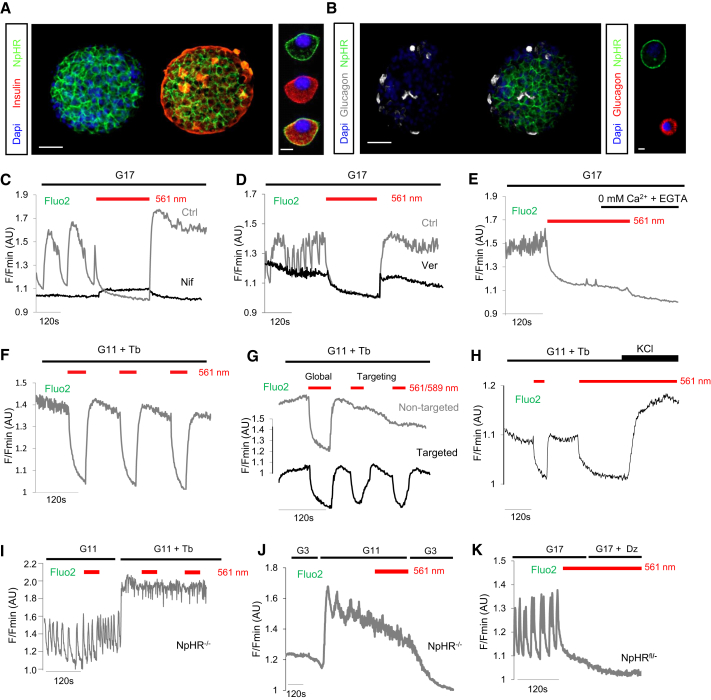
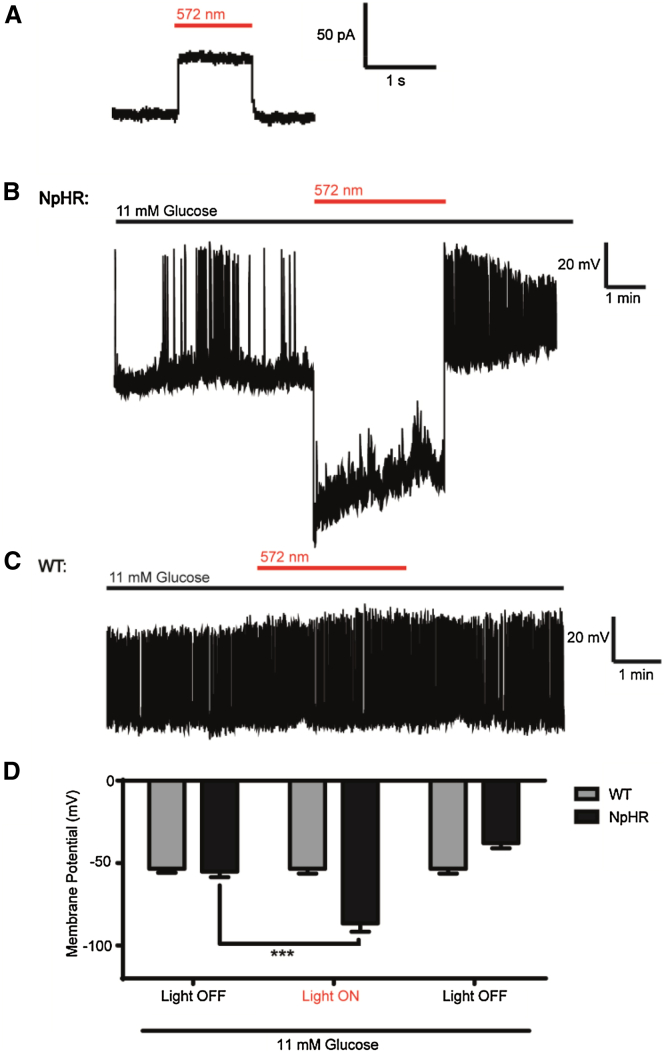
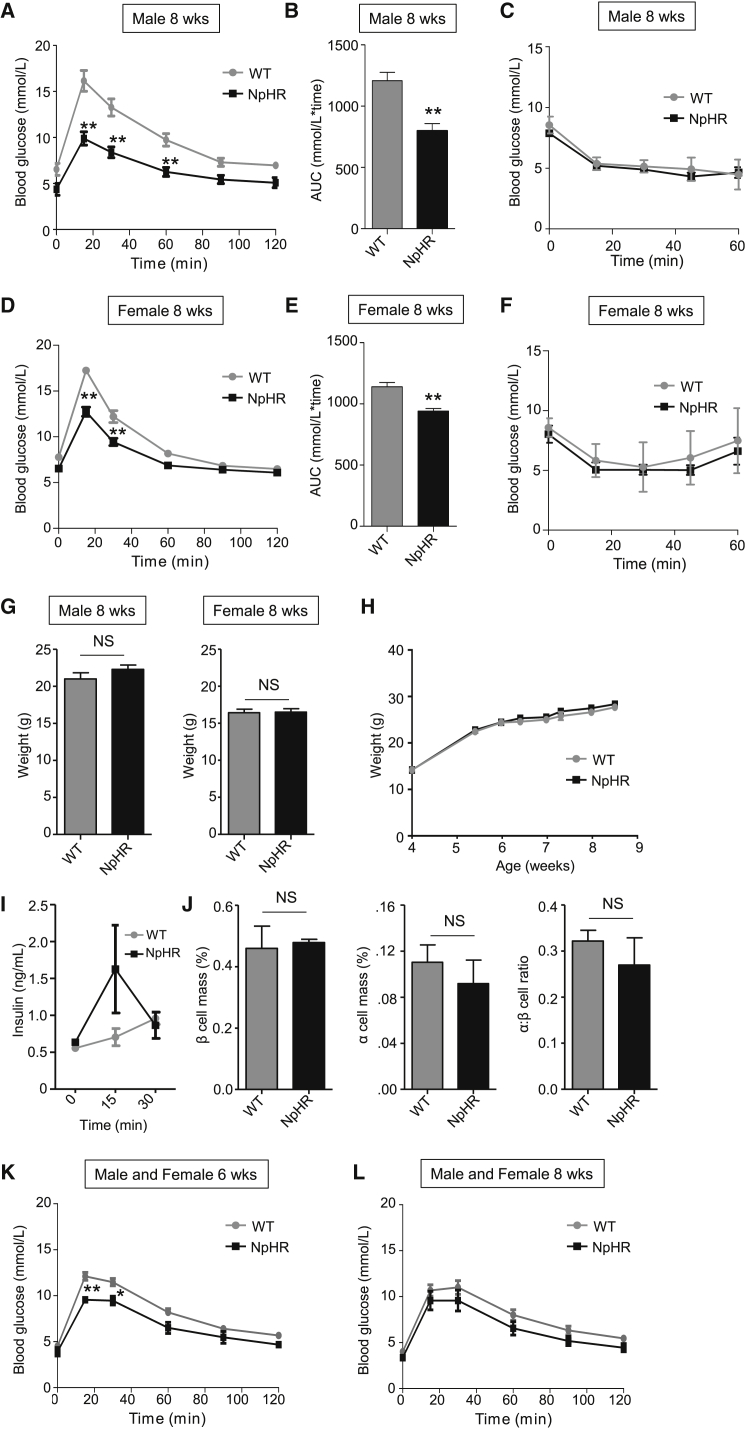
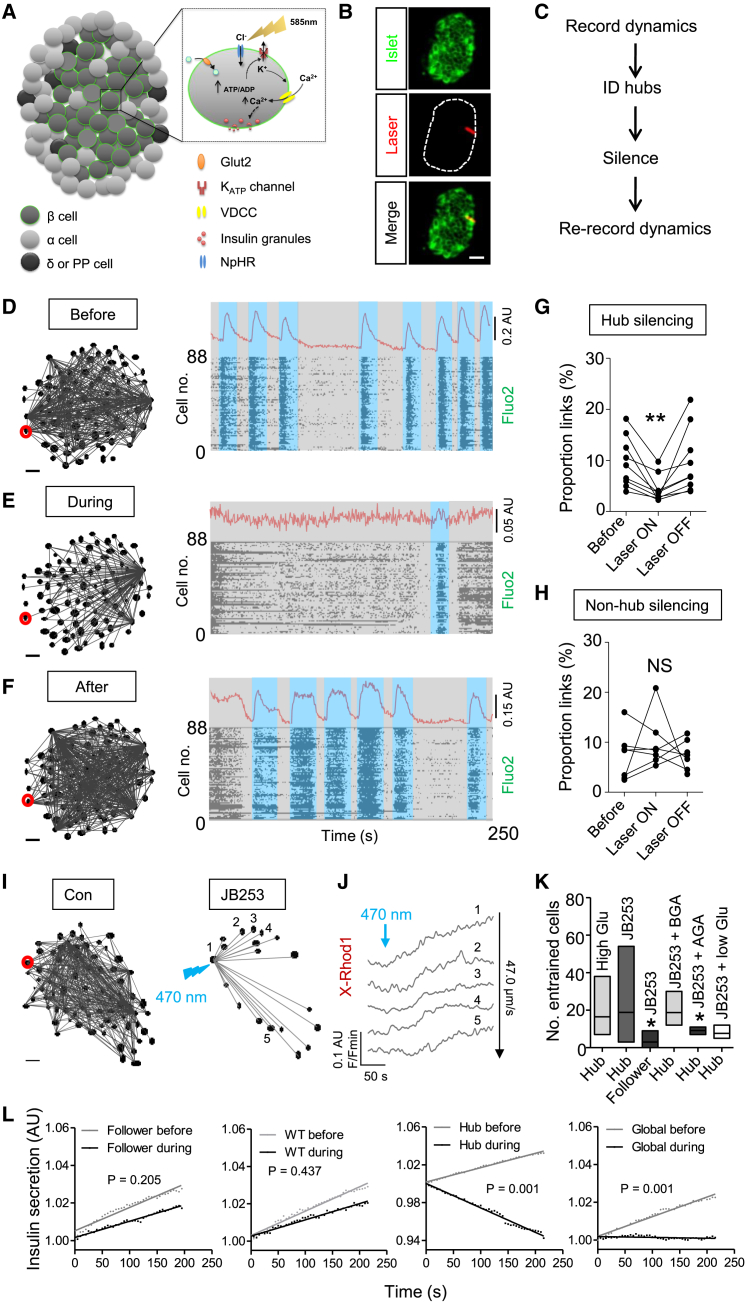
The arrangement of β cells within islets of Langerhans is critical for insulin release through the generation of rhythmic activity. A privileged role for individual β cells in orchestrating these responses has long been suspected, but not directly demonstrated. We show here that the β cell population in situ is operationally heterogeneous. Mapping of islet functional architecture revealed the presence of hub cells with pacemaker properties, which remain stable over recording periods of 2 to 3 hr. Using a dual optogenetic/photopharmacological strategy, silencing of hubs abolished coordinated islet responses to glucose, whereas specific stimulation restored communication patterns. Hubs were metabolically adapted and targeted by both pro-inflammatory and glucolipotoxic insults to induce widespread β cell dysfunction. Thus, the islet is wired by hubs, whose failure may contribute to type 2 diabetes mellitus.
Keywords: diabetes; imaging; insulin; islets; optogenetics; β cells.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures


Comment in
-
Specialized Hub Beta Cells Trade Maximal Insulin Production for Perfect Timing.Cell Metab. 2016 Sep 13;24(3):371-373. doi: 10.1016/j.cmet.2016.08.022. Cell Metab. 2016. PMID: 27626196
-
Metabolic Signaling Drives IFN-γ.Cell Metab. 2016 Nov 8;24(5):651-652. doi: 10.1016/j.cmet.2016.10.018. Cell Metab. 2016. PMID: 27829134 Free PMC article.
References
-
- Ammälä C., Larsson O., Berggren P.-O., Bokvist K., Juntti-Berggren L., Kindmark H., Rorsman P. Inositol trisphosphate-dependent periodic activation of a Ca(2+)-activated K+ conductance in glucose-stimulated pancreatic β-cells. Nature. 1991;353:849–852. - PubMed
-
- Andresen V., Alexander S., Heupel W.M., Hirschberg M., Hoffman R.M., Friedl P. Infrared multiphoton microscopy: subcellular-resolved deep tissue imaging. Curr. Opin. Biotechnol. 2009;20:54–62. - PubMed
MeSH terms
Substances
Grants and funding
- MR/N00275X/1/MRC_/Medical Research Council/United Kingdom
- MR/L02036X/1/MRC_/Medical Research Council/United Kingdom
- MR/K001981/1/MRC_/Medical Research Council/United Kingdom
- MC_PC_14100/MRC_/Medical Research Council/United Kingdom
- BB/J015873/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
