

OPN-a induces muscle inflammation by increasing recruitment and activation of pro-inflammatory macrophages
- PMID: 27452303
- PMCID: PMC5095808
- DOI: 10.1113/EP085768
OPN-a induces muscle inflammation by increasing recruitment and activation of pro-inflammatory macrophages
Abstract
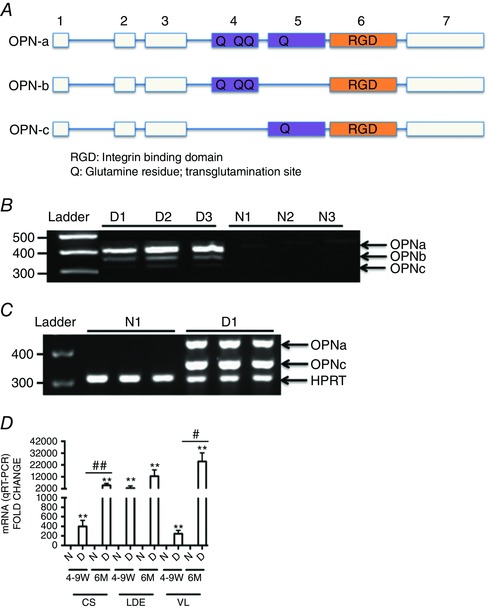
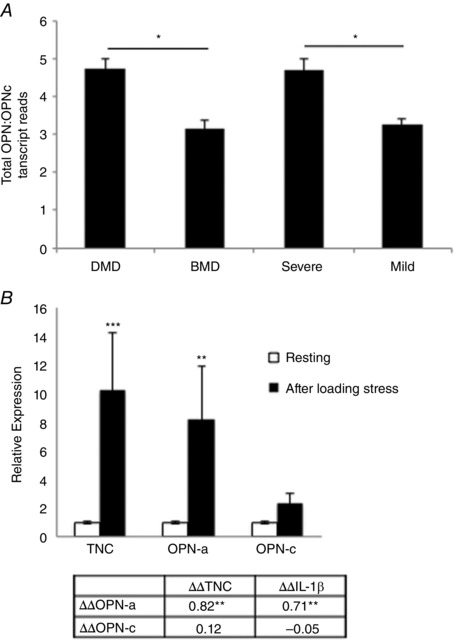
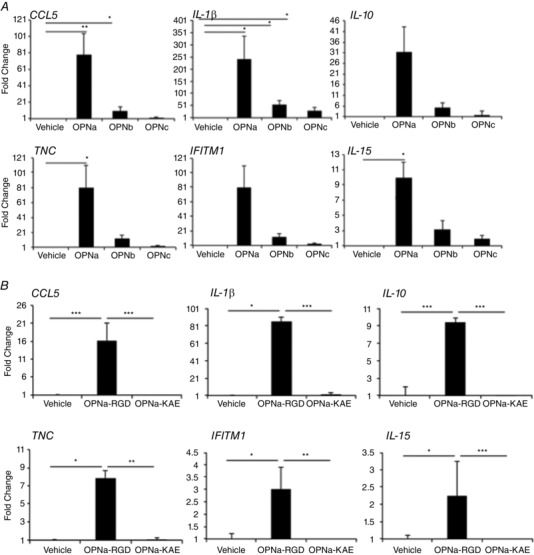
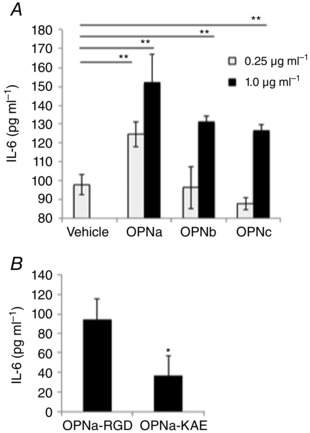
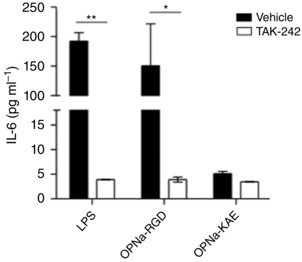
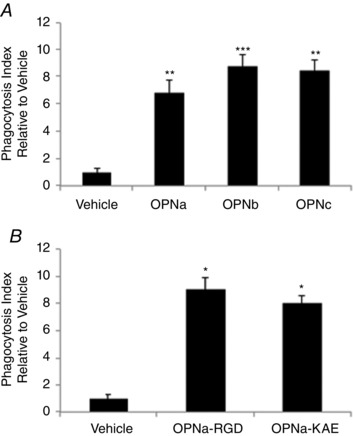
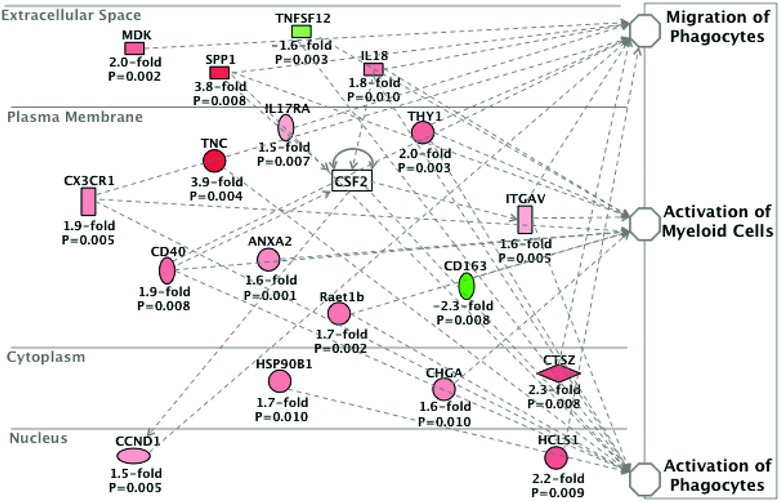
What is the central question of this study? What is the functional relevance of OPN isoform expression in muscle pathology? What is the main finding and its importance? The full-length human OPN-a isoform is the most pro-inflammatory isoform in the muscle microenvironment, acting on macrophages and myoblasts in an RGD-integrin-dependent manner. OPN-a upregulates expression of tenascin-C (TNC), a known Toll-like receptor 4 (TLR4) agonist. Blocking TLR4 signalling inhibits the pro-inflammatory effects of OPN-a, suggesting that a potential mechanism of OPN action is by promoting TNC-TLR4 signalling. Although osteopontin (OPN) is an important mediator of muscle remodelling in health and disease, functional differences in human spliced OPN variants in the muscle microenvironment have not been characterized. We thus sought to define the pro-inflammatory activities of human OPN isoforms (OPN-a, OPN-b and OPN-c) on cells present in regenerating muscle. OPN transcripts were quantified in normal and dystrophic human and dog muscle. Human macrophages and myoblasts were stimulated with recombinant human OPN protein isoforms, and cytokine mRNA and protein induction was assayed. OPN isoforms were greatly increased in dystrophic human (OPN-a > OPN-b > OPN-c) and dog muscle (OPN-a = OPN-c). In healthy human muscle, mechanical loading also upregulated OPN-a expression (eightfold; P < 0.01), but did not significantly upregulate OPN-c expression (twofold; P > 0.05). In vitro, OPN-a displayed the most pronounced pro-inflammatory activity among isoforms, acting on both macrophages and myoblasts. In vitro and in vivo data revealed that OPN-a upregulated tenascin-C (TNC), a known Toll-like receptor 4 (TLR4) agonist. Inhibition of TLR4 signalling attenuated OPN-mediated macrophage cytokine production. In summary, OPN-a is the most abundant and functionally active human spliced isoform in the skeletal muscle microenvironment. Here, OPN-a promotes pro-inflammatory signalling in both macrophages and myoblasts, possibly through induction of TNC-TLR4 signalling. Together, our findings suggest that specific targeting of OPN-a and/or TNC signalling in the damaged muscle microenvironment may be of therapeutic relevance.
Keywords: Duchenne muscular dystrophy; OPN-a; TNC; muscle inflammation; osteopontin; skeletal muscle.
© 2016 The Authors. Experimental Physiology published by John Wiley & Sons Ltd on behalf of The Physiological Society.
Figures
References
-
- Bandopadhyay M, Bulbule A, Butti R, Chakraborty G, Ghorpade P, Ghosh P, Gorain M, Kale S, Kumar D, Kumar S, Totakura KV, Roy G, Sharma P, Shetti D, Soundararajan G, Thorat D, Tomar D, Nalukurthi R, Raja R, Mishra R, Yadav AS & Kundu GC (2014). Osteopontin as a therapeutic target for cancer. Expert Opin Ther Targets 18, 883–895. - PubMed
-
- Bello L, Kesari A, Gordish‐Dressman H, Cnaan A, Morgenroth LP, Punetha J, Duong T, Henricson EK, Pegoraro E, McDonald CM, Hoffman EP & Cooperative International Neuromuscular Research Group Investigators (2015). Genetic modifiers of ambulation in the Cooperative International Neuromuscular Research Group Duchenne Natural History Study. Ann Neurol 77, 684–696. - PMC - PubMed
-
- Cavaillon JM & Haeffner‐Cavaillon N (1986). Polymyxin‐B inhibition of LPS‐induced interleukin‐1 secretion by human monocytes is dependent upon the LPS origin. Mol Immunol 23, 965–969. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous