

Structure and Function Analysis of an Antibody Recognizing All Influenza A Subtypes
- PMID: 27453466
- PMCID: PMC4967455
- DOI: 10.1016/j.cell.2016.05.073
Structure and Function Analysis of an Antibody Recognizing All Influenza A Subtypes
Abstract
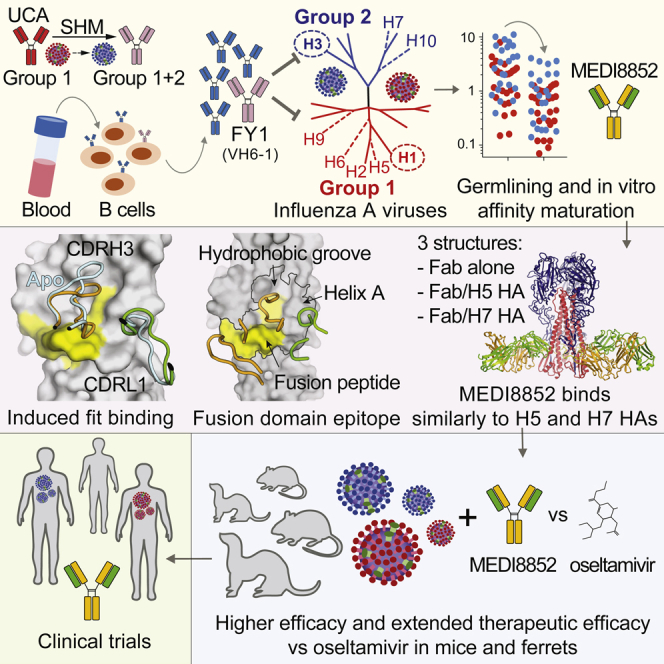
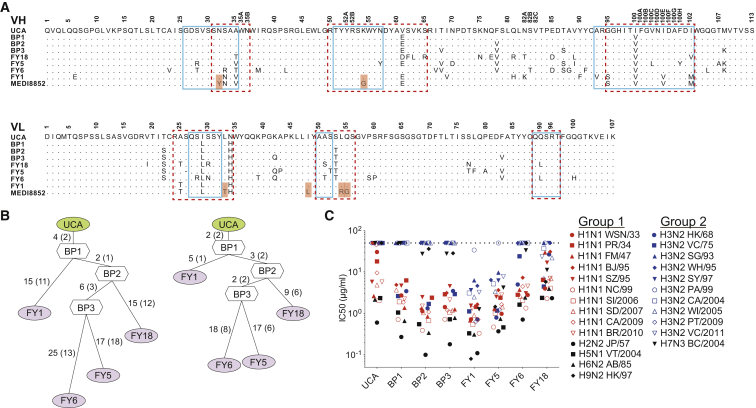
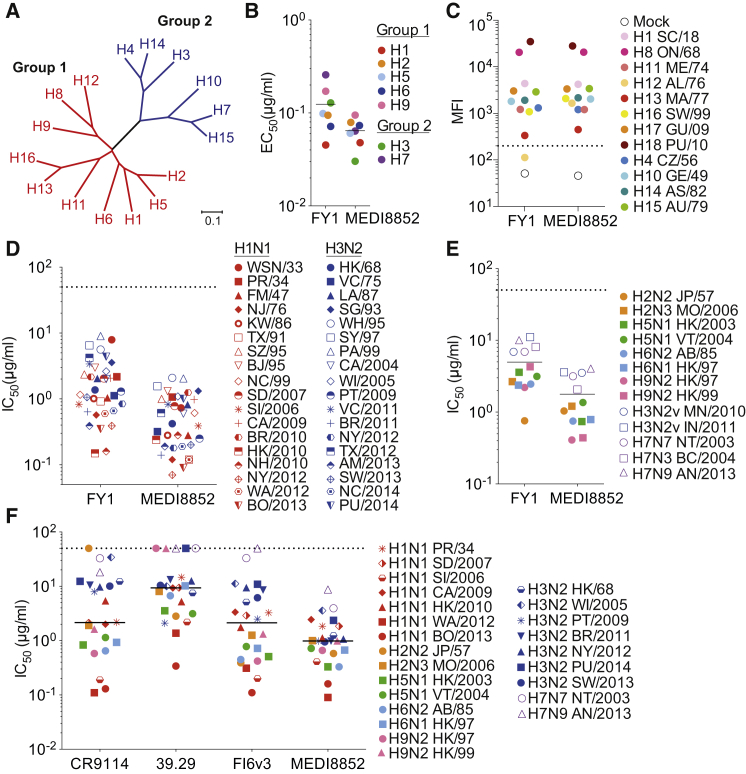
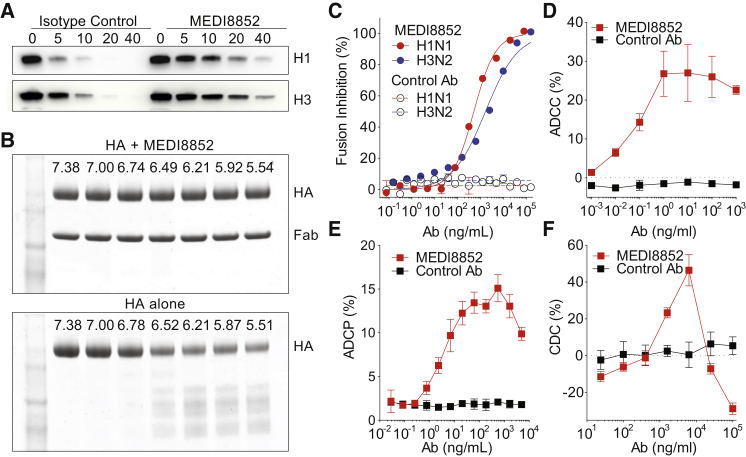
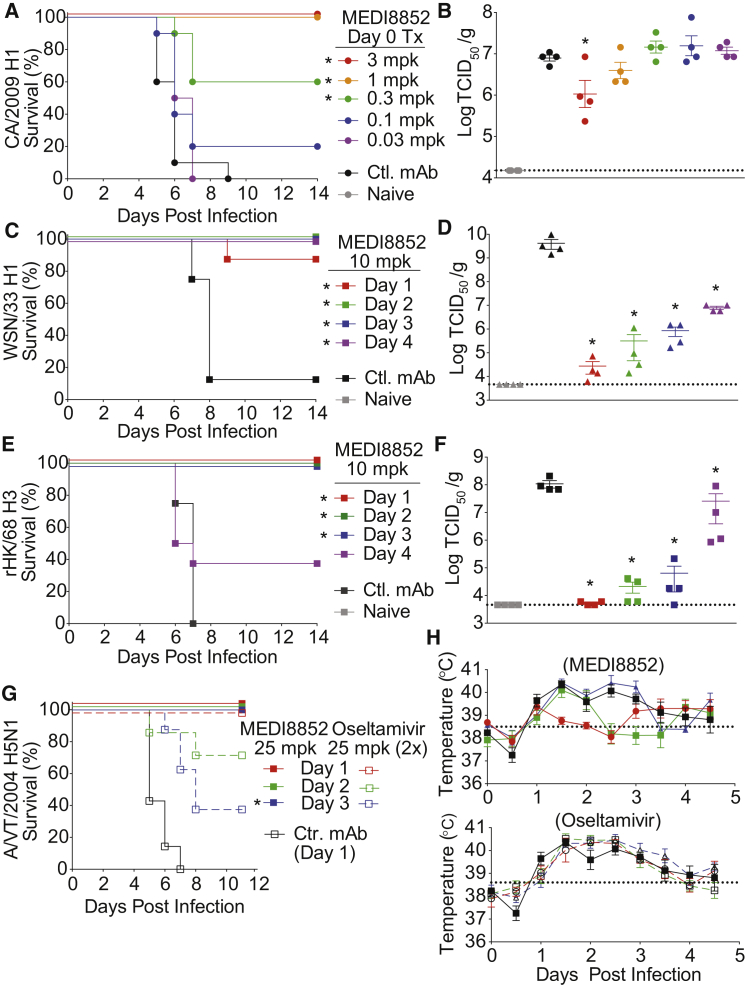
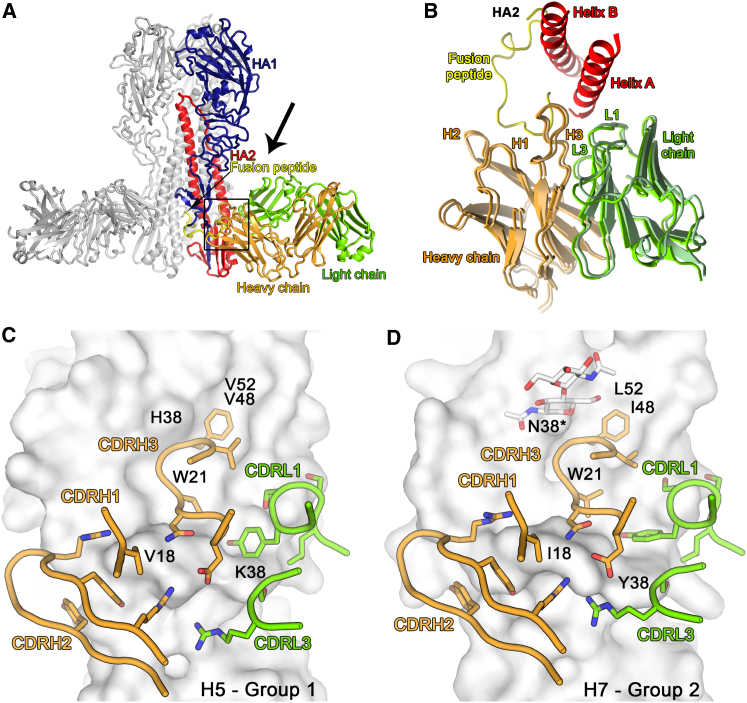
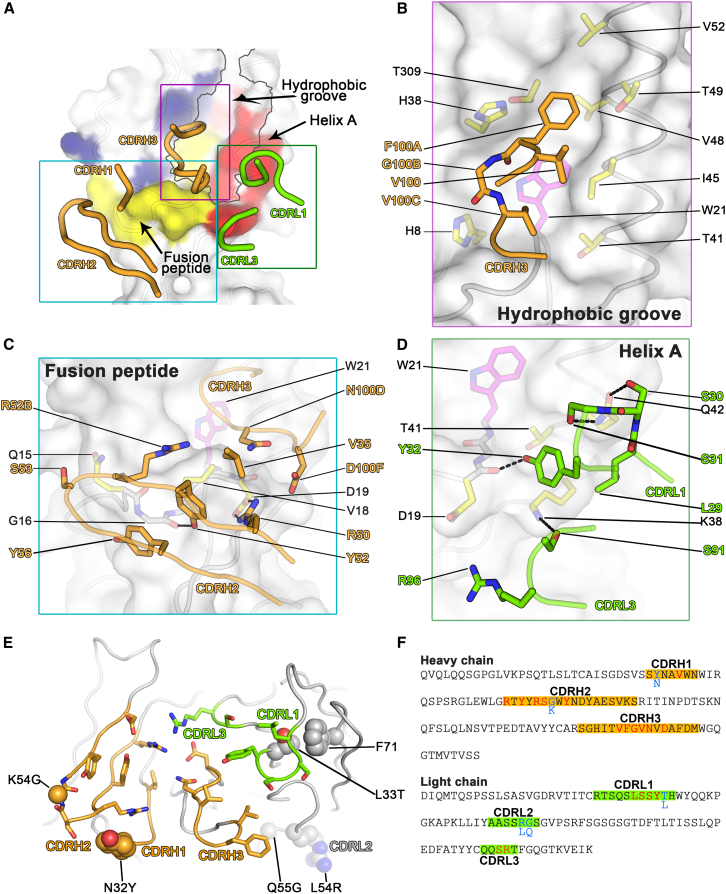
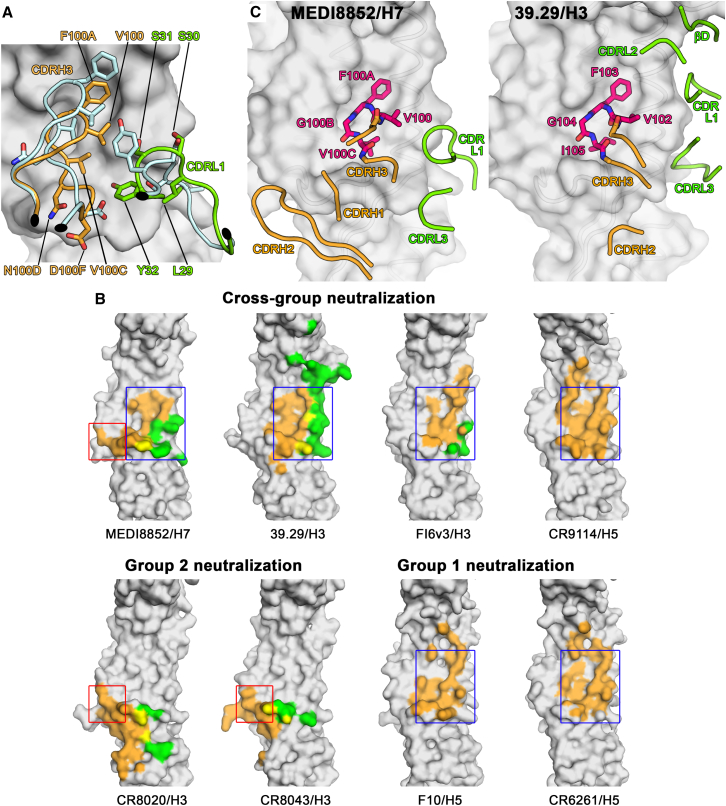
Influenza virus remains a threat because of its ability to evade vaccine-induced immune responses due to antigenic drift. Here, we describe the isolation, evolution, and structure of a broad-spectrum human monoclonal antibody (mAb), MEDI8852, effectively reacting with all influenza A hemagglutinin (HA) subtypes. MEDI8852 uses the heavy-chain VH6-1 gene and has higher potency and breadth when compared to other anti-stem antibodies. MEDI8852 is effective in mice and ferrets with a therapeutic window superior to that of oseltamivir. Crystallographic analysis of Fab alone or in complex with H5 or H7 HA proteins reveals that MEDI8852 binds through a coordinated movement of CDRs to a highly conserved epitope encompassing a hydrophobic groove in the fusion domain and a large portion of the fusion peptide, distinguishing it from other structurally characterized cross-reactive antibodies. The unprecedented breadth and potency of neutralization by MEDI8852 support its development as immunotherapy for influenza virus-infected humans.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
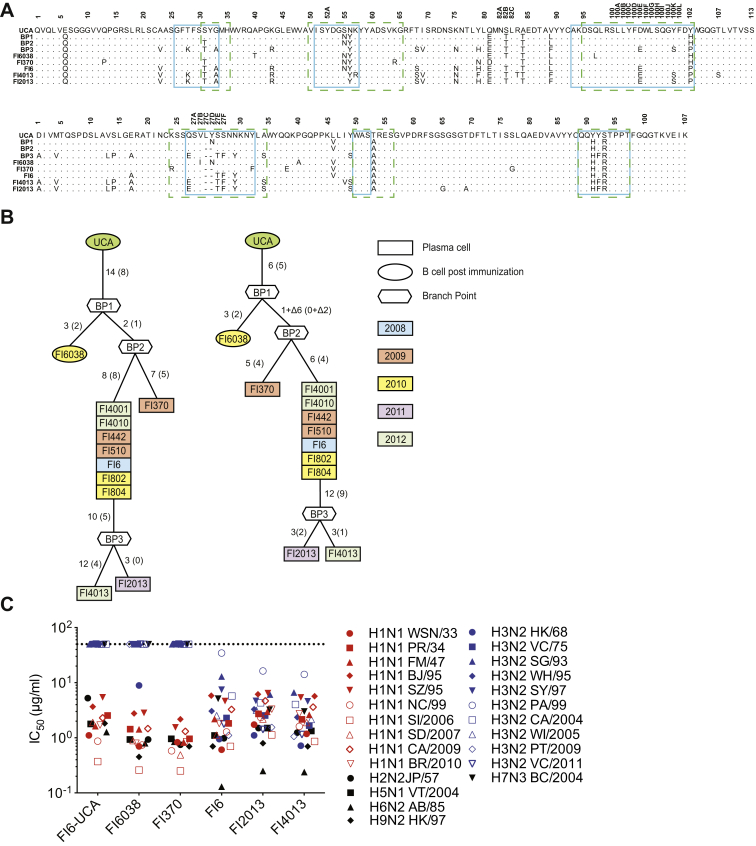
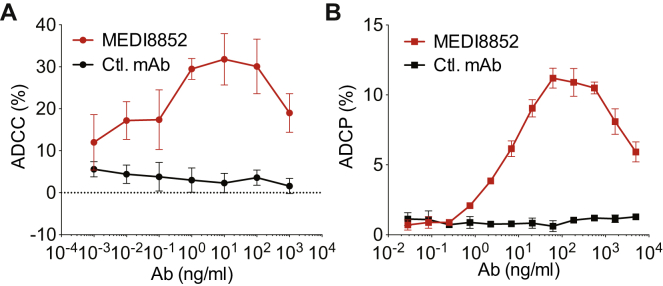
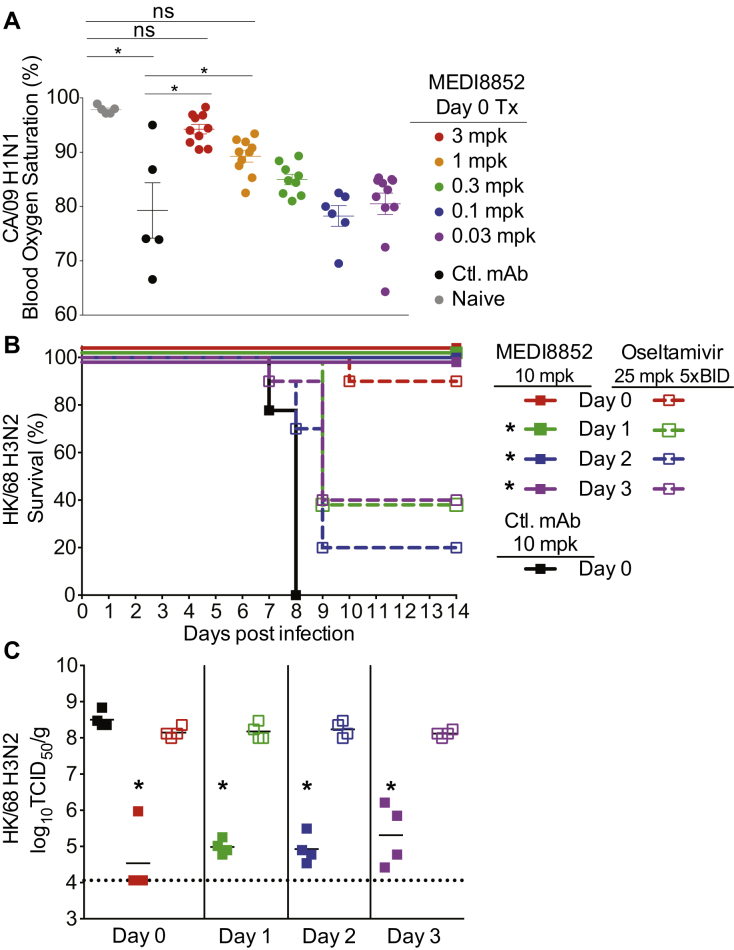
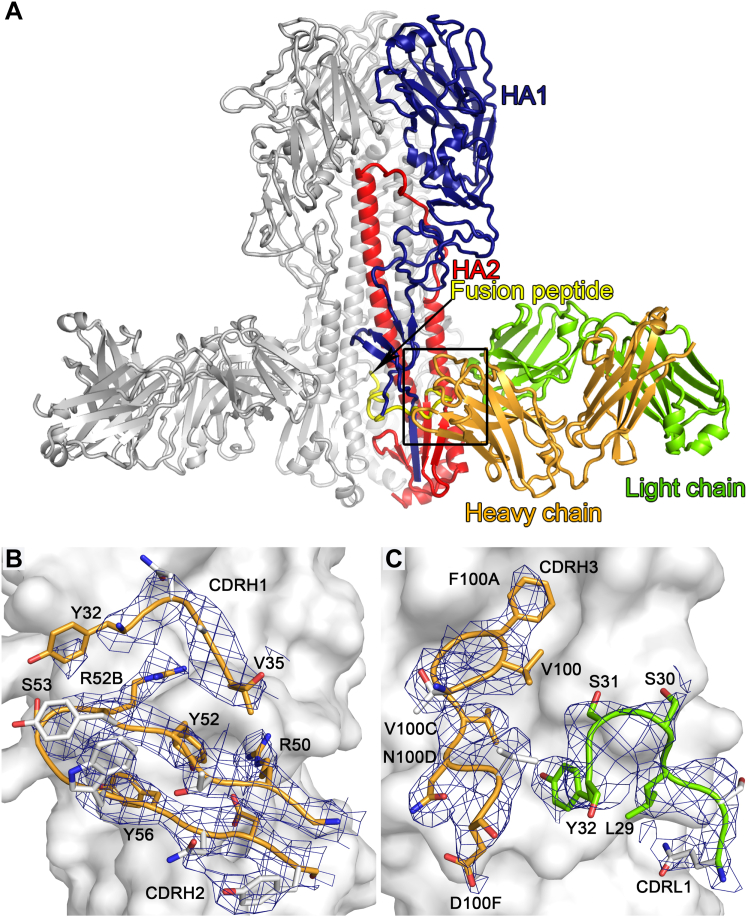
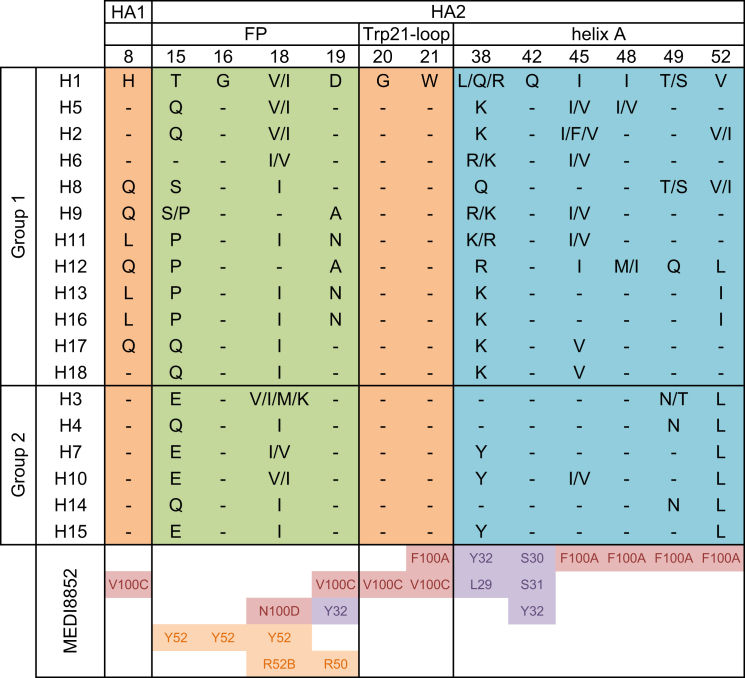
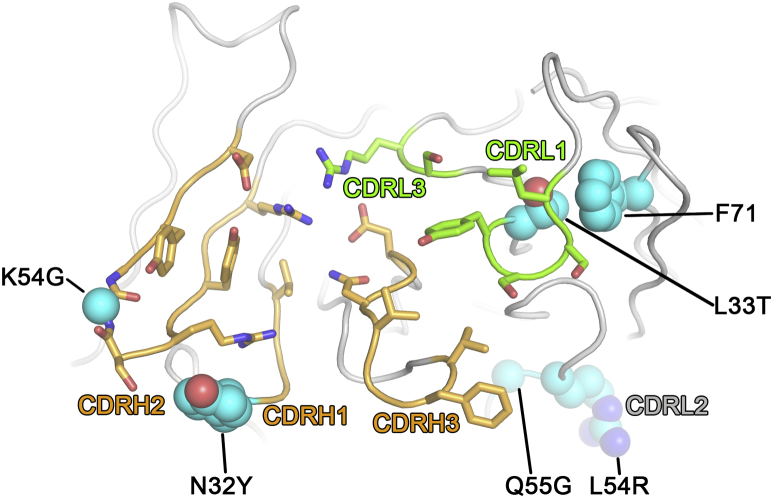
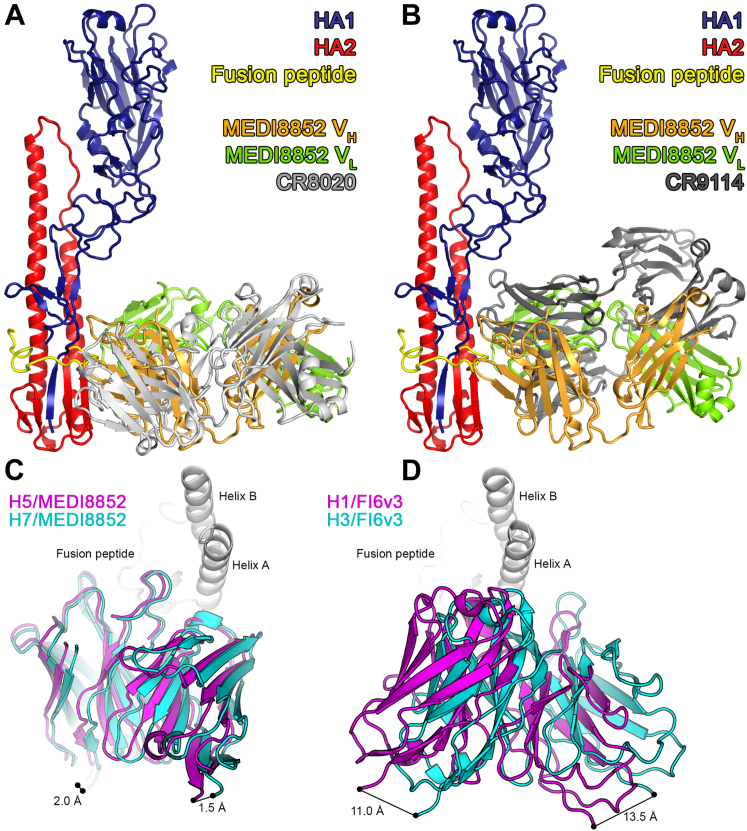
Comment in
-
Broadening Horizons: New Antibodies Against Influenza.Cell. 2016 Jul 28;166(3):532-533. doi: 10.1016/j.cell.2016.07.023. Cell. 2016. PMID: 27471961
References
-
- Benjamin E., Wang W., McAuliffe J.M., Palmer-Hill F.J., Kallewaard N.L., Chen Z., Suzich J.A., Blair W.S., Jin H., Zhu Q. A broadly neutralizing human monoclonal antibody directed against a novel conserved epitope on the influenza virus H3 hemagglutinin globular head. J. Virol. 2014;88:6743–6750. - PMC - PubMed
-
- Corti D., Lanzavecchia A. Broadly neutralizing antiviral antibodies. Annu. Rev. Immunol. 2013;31:705–742. - PubMed
-
- Corti D., Suguitan A.L., Jr., Pinna D., Silacci C., Fernandez-Rodriguez B.M., Vanzetta F., Santos C., Luke C.J., Torres-Velez F.J., Temperton N.J. Heterosubtypic neutralizing antibodies are produced by individuals immunized with a seasonal influenza vaccine. J. Clin. Invest. 2010;120:1663–1673. - PMC - PubMed
-
- Corti D., Voss J., Gamblin S.J., Codoni G., Macagno A., Jarrossay D., Vachieri S.G., Pinna D., Minola A., Vanzetta F. A neutralizing antibody selected from plasma cells that binds to group 1 and group 2 influenza A hemagglutinins. Science. 2011;333:850–856. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
